Подвиды
Широко распространенные виды состоят из практически бесконечного числа локальных популяций, каждая из которых объединяет генетически различных особей. Совокупность локальных популяций, населяющих определенную часть ареала вида и таксономически, т. е. по ряду внешних диагностических признаков, отличающихся от других популяций этого вида, называется подвидом (до сих пор иногда в качестве синонима используется термин "географическая раса"). Важно помнить, что подвид обычно состоит из многих локальных популяций, видимо, всегда хотя бы слегка отличающихся друг от друга генетически и фенотипически. Как правило, каждую локальную популяцию удается отнести к тому или иному подвиду, но для каждой отдельной особи это не всегда удается из-за индивидуальной изменчивости, иногда совпадающей с изменчивостью соседних подвидов. Поэтому при выделении подвидов большинство систематиков сейчас придерживается "правила 75%": подвидом следует считать популяцию или группу популяций, не менее 75% особей которой отличимы от всех особей другого (ближайшего) подвида. Чем больше серии экземпляров, с которыми работает систематик, тем больше шансов выявить истинное положение.
Подвид занимает определенную территорию; два подвида не могут встречаться на одной и той же территории. Если эти популяции свободно скрещиваются, то данные "подвиды" выделены произвольно и требуют пересмотра. Если же между этими популяциями существует репродуктивная изоляция, то это самостоятельные виды. Между подвидами одного вида репродуктивной изоляции быть не может (немногие исключения в сложных таксономических случаях приведены ниже; см. кольцевые перекрывания). Таким образом подвиды всегда аллопатричны, т. е. географически исключают друг друга, но обычно занимают смежные (соседние) территории. Подвиды более или менее постепенно переходят один в другой, образуя зону интерградации (перекрывания), где между двумя подвидами идет свободное скрещивание и многие особи имеют промежуточные признаки. Выявление границ подвидов - дело трудное, а сами границы в некоторой степени условны.
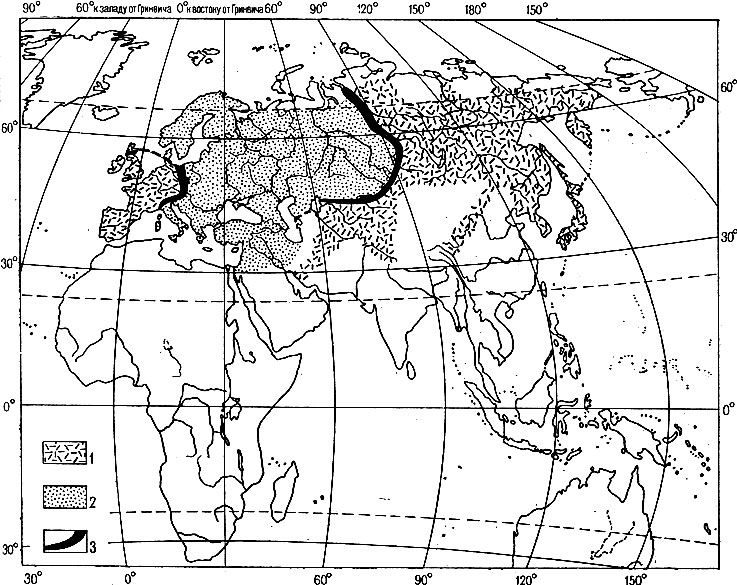
Рис. 3. Зоны гибридизации между серыми и черными воронами в Палеарктике (по Майзе, 1928; упрощено); 1 - ареал черных ворон группы corone, 2 - ареал серых ворон группы cornix, 3 - зона гибридизации
В некоторых случаях, при резких подвидовых отличиях, пограничная зона интерградации между подвидами выражена четко. Примером могут служить западные черный и серый подвиды обыкновенной вороны (Corvus соrоnе соrоnе и Corvus corone comix), контактирующие в Западной Европе в узкой по сравнению с ареалами этих подвидов полосе шириной в 50 - 250 км (рис. 3). Спаривание (гибридизация) в зоне интерградации идет беспорядочно и здесь встречаются все возможные комбинации родительских и промежуточных признаков. Такая же зона гибридизации (интерградации) между восточными подвидами вороны (серой Corvus соrоnе sharpii и черной Corvus соronе orientalis) проходит по казахстанским степям, северо-западному Алтаю, Прибалхашью и по долине Енисея. Полагают, что в период наибольшего оледенения ареал вороны был разделен на участки, где живущие в изоляции популяции образовали подвиды, резко разошедшиеся по морфологическим признакам, прежде всего по окраске и соответствующим генным комплексам. После отступания льдов, в послеледниковый период эти резко различающиеся внешне, но не выработавшие репродуктивной изоляции подвиды стали расселяться, образовав в районах стыков своих ареалов гибридные зоны. Относительную узость и устойчивость гибридных зон, видимо, можно объяснить тем, что отбор отбрасывает дисгармоничные комбинации генных комплексов в зоне гибридизации и отбор же препятствует распространению этих промежуточных комбинаций в смежные "чистые" подвидовые популяции (Майр, 1968). Многие авторы такие географические формы с четкими узкими зонами гибридизации рассматривают как самостоятельные виды; часть систематиков считает самостоятельными видами черную и серую ворон, что кажется не вполне обоснованным и явно противоречит широкой концепции биологического вида.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'