Происхождение и эволюция класса птиц
Палеонтологические материалы по птицам, как уже указывалось, очень скудны и отрывочны. Поэтому говорить о предках птиц среди рептилий и о происхождении и взаимоотношениях современных групп птиц приходится преимущественно предположительно, основываясь на морфологических особенностях современных групп и, в очень малой степени, на отрывочных палеонтологических данных.
По современным представлениям, предками птиц были архозавры Аrchosauria - обширная и очень разнообразная группа рептилий, господствовавшая в мезозое и включавшая различных динозавров, крокодилов, летающих ящеров. Непосредственных предков птиц, видимо, нужно искать среди наиболее древней и примитивной группы архозавров - текодонтов или псевдозухий Thecodontia (Pseudosuchia), давших начало и остальным более высокоорганизованным группам архозавров. Псевдозухии по внешнему виду были похожи на ящериц. Они вели преимущественно наземный образ жизни, питались, видимо, разнообразной мелкой животной пищей. Задние ноги были несколько длиннее передних, и при быстром беге животные опирались о почву только задними конечностями. По многим морфологическим особенностям псевдозухии весьма сходны с птицами (детали строения черепа, таза, задних конечностей). Промежуточных форм, которые позволяли бы выяснить стадии развития оперения и выработку способности к полету, пока не найдено. Полагают, что переход к древесному образу жизни и приспособление к прыжкам с ветки на ветку сопровождался разрастанием и усложнением структуры роговых чешуй на боках туловища и хвоста, по заднему краю передних и задних конечностей, что обеспечивало возможность планирования и удлинения прыжков (гипотетическая стадия предптицы, по Хейльманну, 1926, рис. 13).
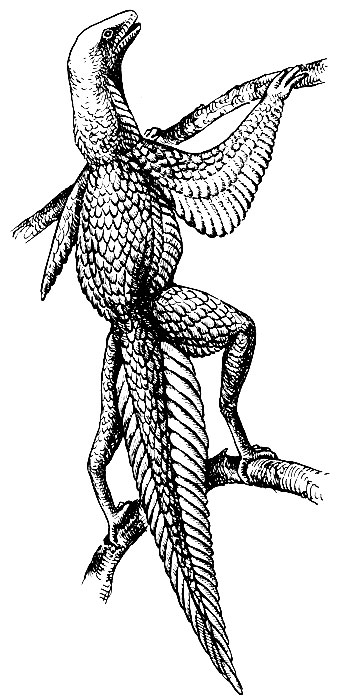
Рис. 13. Гипотетический предок птиц - предптица (по Хейльманну, 1926)
Дальнейшая специализация и отбор привели к развитию оперения крыла, обеспечившего возможность сначала планирующего, а затем активного полета. Оперение, покрывающее все тело птиц, вероятно, развивалось не столько как приспособление к полету (обеспечение обтекаемости тела), сколько как приспособление к термоизоляции; высказываются предположения, что оно могло сформироваться еще до выработки способности к полету. У некоторых псевдозухий были удлиненные роговые чешуи с четким продольным гребнем, от которого отходили мелкие поперечные ребрышки. Такие чешуйки, видимо, можно рассматривать как структуру, из которой путем расчленения могло возникнуть птичье перо.
Обособление птиц от пресмыкающихся, видимо, произошло уже в конце триаса - начале юры (170 - 190 млн. лет назад). Однако ископаемых остатков птиц этого времени пока не найдено. Довольно хорошие остатки двух самых древних известных нам птиц были найдены в 60-х годах XIX в. у Золенгофена (Бавария) на разработках сланцевых песчаников, представляющих отложения мелководного залива юрского моря (возраст около 150 млн. лет). Их назвали Archaeopteryx lithographica и Archaeornis simetisi. В 1956 г. там же (в радиусе до 300 ж, но на иной глубине) найден значительно худшей сохранности отпечаток еще одной особи. Часть исследователей рассматривает эти находки как остатки трех разных видов (два вида рода Archaeopteryx и один рода Archaeornis), другие же считают представителями одного вида Archaeopteryx lithographica, объясняя различия между особями как индивидуальные, возрастные и половые. Вопрос этот нуждается в дополнительном изучении, но и сейчас совершенно очевидна очень большая близость всех этих особей. Все они имеют хорошо развитое оперение крыльев, хвоста и тела, некоторые пропорции черепа ближе к птичьим, чем к рептильным, птичьи черты несут пояс передних конечностей и таз, задние конечности. Типично рептильные черты: нет рогового клюва, имеются слабые зубы, кисть несет первостепенные маховые, но пряжка еще не образовалась и есть три хорошо развитых подвижных пальца с крупными когтями, образование сложного крестца только началось (позвонки еще не срастаются, подвижные кости соединяются с небольшим числом позвонков), бедро причленяется под тупым углом (направлено несколько вбок), проксимальные тарзальные кости еще не приросли к концу большой берцовой кости, хорошо развита малая берцовая кость, не завершено образование цевки и т. п. По внешнему облику, ряду внутренних морфологических признаков и, насколько это можно предполагать сейчас, образу жизни архептерикс (и археорнис) - древние, примитивные, но птицы.
Подкласс настоящих, или веерохвостых, птиц Neornithes, видимо, - прямые потомки каких-то ящерохвостых птиц Archaeornithes. Некоторые исследователи (например, Гадов) считают за прямых предков птиц археоптерикса и археорниса, другие (их, пожалуй, большинство) рассматривают группу археоптерикса как примитивную, но специализированную боковую и слепую ветвь, полагая, что предками современных птиц должны быть другие, пока еще не обнаруженные ящерохвостые птицы (в случае их обнаружения они, вероятно, образуют другой отряд этого подкласса).
Происхождение и взаимоотношения современных групп (отрядов) птиц остаются, как уже говорилось, за отсутствием достаточных палеонтологических материалов, как правило, очень гипотетическими. Наибольшее внимание уделил этому вопросу М. Фюрбрингер, заметно меньшее - Г. Гадов. Не внесли много нового и бесспорного в эту проблему и современные авторы, в той или иной степени касавшиеся филогении птиц. Поэтому здесь изложено представление о взаимоотношениях отдельных групп птиц преимущественно по Фюрбрингеру, но в ряде случаев, где это нужно, внесены некоторые изменения (в частных случаях эти изменения специально не оговариваются).
Ископаемые остатки веерохвостых птиц обнаружены только в меловом периоде (их возраст примерно 80 - 90 млн. лет), т. е. они на несколько десятков миллионов лет моложе археоптериксов. По нескольким плохой сохранности остаткам описаны гесперорнисы - очень крупные (длиной более 100 см) птицы, внешне, видимо, несколько похожие на гагар или поганок, совершенно не летавшие (грудина без киля, от передней конечности сохранилось только маленькое тоненькое плечо), но, вероятно, они хорошо плавали и ныряли: челюсти их несли зубы. В меловых же отложениях найдены остатки других зубастых птиц - ихтиорнисов. Они имели хорошо развитый киль грудины, типичное птичье крыло и, видимо, обладали активным полетом. По размерам они были примерно с голубя. Родственные отношения зубастых меловых птиц с другими птицами очень неясны. Возможно, часть этих остатков относится к меловым динозаврам, а не птицам, но часть, несомненно, зубастые птицы. Фюрбрингер сближал гесперорнисов с гагарами и поганками, а их тиорнисов - с чайками, но при этом подчеркивал их очень давнюю обособленность.
Гадов гесперорнисов выделял в надотряд, показывая этим их обособленное положение, а ихтиорнисы как самостоятельный отряд (тоже обособленный) начинали перечень отрядов килегрудных птиц. Подчеркивая неясность происхождения (да и объем) этих групп, Уэтмор (1960) выделяет их в самостоятельные надотряды.
Из меловых отложений описано еще несколько семейств птиц (по обломкам челюстей и трубчатых костей), но их положение и внешний облик очень неясны: возможно, хотя бы часть этих находок - остатки рептилий. Встреченные в отложениях третичного периода кайнозойской эры (возраст менее 55 млн. лет) остатки птиц с большей или меньшей вероятностью могут быть отнесены к современным отрядам. Во всяком случае они принадлежат к типичным птицам, близким к тем или иным группам современных птиц. Судя по этим материалам, можно предполагать, что интенсивная адаптивная радиация птиц и становление большинства современных отрядов проходило в самом конце мелового периода мезозойской эры - в третичный период кайнозойской эры, т. е. в период примерно 70 - 40 млн. лет назад.
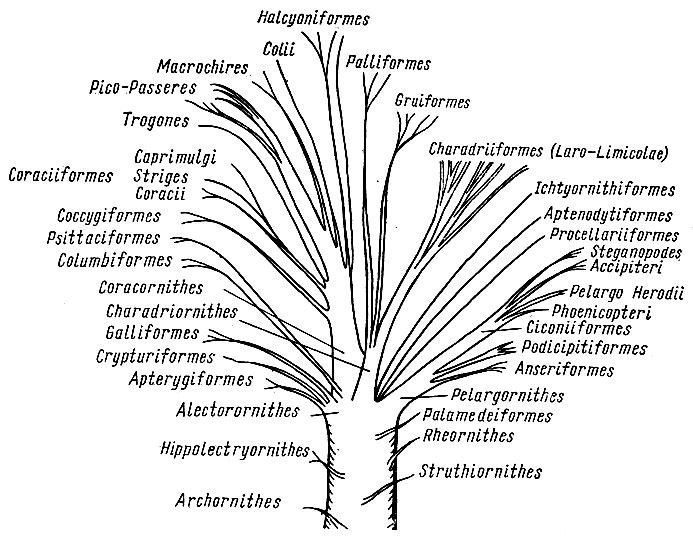
Рис. 14. Филогенетические отношения в классе птиц (по Фюрбрингеру, 1888; упрощено)
Гадов всех ныне живущих и ископаемых крупных нелетающих страусоподобных птиц выделял в надотряд бескилевых, подчеркивая этим их сходство и филогенетическое родство (т. е. монофилию), а также их определенную обособленность от остальных групп птиц. Среди килевых птиц наиболее близки к страусоподобным тинаму (скрытохвосты), куриные, журавлеобразные и близкие к ним группы. Фюрбрингер и большинство современных авторов (Уэтмор, Штреземанн и др.) считают страусоподобных птиц сборной полифилетической группой, разбиваемой на ряд самостоятельных отрядов, начинающих систему, что подразумевает их определенную примитивность и изолированность. Их общие сходные черты - результат конвергенции: крупные размеры, потеря способности к полету, приспособление к быстрому бегу и т. д.
По Гадову, эволюция остальных птиц шла в двух направлениях (две ветви, см. выше), каждое из которых в свою очередь разделяется на 2 группы родственных отрядов. Поганко-гагарообразные (ихтиорнисы, поганки, гагары, пингвины и трубконосые) родственны группе Pelargomorphae (аистообразные, гусеобразные, хищники), а близкая к страусоподобным группа курообразных (тинаму, куриные, журавлеобразные, куликообразные) дала начало ракшеобразным (кукушки, ракши, воробьиные).
Эволюционные представления Фюрбрингера в общих чертах близки к этой схеме Гадова (о иных взглядах на страусоподобных говорилось выше), но он излагает свои представления с большей степенью детализации. Крупный ствол птиц - отряд Pelargornithes - делится на ряд ветвей: гусеобразных, поганкообразных (включая поганок, гагар, гесперорнисов); от основания этих ветвей отходит еще одна мощная ветвь, которая, разделяясь, дает начало веслоногим, дневным хищникам и группам подотряда аистообразных (рис. 14).
Отряд паламедей Palamedeiformes занимает промежуточное положение между нанду и отрядом Pelargornithes. Промежуточные отряды пингвинов Aptenodytiformes и трубконосых Procellariiformes довольно обособлены; возможно, они имеют давние слабые связи друг с другом и, еще более отдаленные, с отрядами Pelargornithes и куликообразных Charadriornithes. Ихтиорнисы Ichthyornithiformes - тоже промежуточный отряд, но с несколько более выраженными давними связями с куликообразными. Отряд куликообразных Charadriornithes очень давними связями видимо может быть приближен к отряду ракшеобразных. Основные ветвления ствола куликообразных - подотряд Laro-Limicolae - кулики, чайки и чистики; к ним довольно близки зобатые бегунки и дрофы (переходной группой между куликами и дрофами служат авдотки). От основания ствола куликообразных обособились еще две крупные ветви: журавлеобразные Gruiformes и пастушковые Ralliformes. Журавлеобразные разделяются на несколько ветвей: собственно журавлиных (журавли, арамиды, трубачи, кариамы) и занимающих несколько обособленное положение солнечных цапель и кагу. Пастушковые подразделяются на родственные ветви: пастушков, лапчатоногих и стоящих несколько обособленно трехперсток и мадагаскарских пастушков.
Один из древних стволов родословного древа птиц - отряд курообраз- ных Alectorornithis. Наиболее примитивная ветвь - подотряд бескрылых, или киви Apterygiformes, включающий киви и моа; обе группы разошлись почти сразу после своего обособления. Общим происхождением с предыдущей ветвью, видимо, связаны тинаму (или скрытохвосты) Crypturiformes, в свою очередь через предков близкие к подотряду куриных Galliformes (практически в современном объеме). Отряд курообразных очень давними родственными связями, вероятно, более или менее близок к пастушковым Ralliformes, а через них и ко всему отряду куликообразных.
Промежуточные отряды голубеобразных Columbiformes (включающий рябков и голубей) и попугаев Psittaciformes занимают в системе обособленное положение. Вероятно, через очень давние отношения они как-то связаны с обширным отрядом ракшеобразных. По представлениям Гадова, голуби и рябки в ранге подотрядов входят в отряд куликообразных, а попугаи - тоже как подотряд - в отряд кукушкообразных. По сути дела сходной точки зрения придерживается и Уэтмор, ставящий отряд голубеобразных сразу же за отрядом куликообразных, а отряд попугаев - рядом с отрядом кукушек.
Обширный и разнообразный отряд Coracornithes своими истоками связан с предками отряда куликообразных. Основная и наиболее мощная ветвь - подотряд дятлов и воробьиных Pico - Passeriformes, которая затем разделяется на отдельные группы собственно воробьиных и дятловых. От основания этой ветви обособились Macrochires (стрижи, колибри) и птицы мыши Colii. Еще раньше от этой ветви обособляются трогоны Тгоgones. От самого основания ветви дятлово-воробьиных птиц обособляются и три других подотряда ракшеобразных. Подотряд кукушкообразных Coccygiformes отделяет ветвь якамар Galbulae (якамары и пуховки, а по Уэтмору -это подотряд отряда дятлов), а затем разделяется на кукушек Cuculidae и бананоедов Musophagidae. Подотряд ракшеобразных Со- raciiformes разделяется на собственно ракш Coraciae и на вторую ветвь, вскоре делящуюся на сов Striges и козодоев Caprimulgi. Подотряд зимород- ковых Halcyoniformes (он, видимо, ближе к Pico - Passeriformes, чем к другим подотрядам), отделяя в самом начале боковую веточку тоди Todi (кроме тоди Фюрбрингер сюда включает и момотов), разделяется на 3 родственные группы: зимородковых Halcyones (Alcedinidae), удодов Bucero tes (удоды, птицы-носороги) и щурок Meropes.
Сопоставление филогенетических представлений Фюрбрингера со взглядами Гадова и Уэтмора (очередность и порядок перечисления отрядов) показывает значительное их совпадение; резкие отличия в положении и взаимоотношении отдельных групп указаны выше.
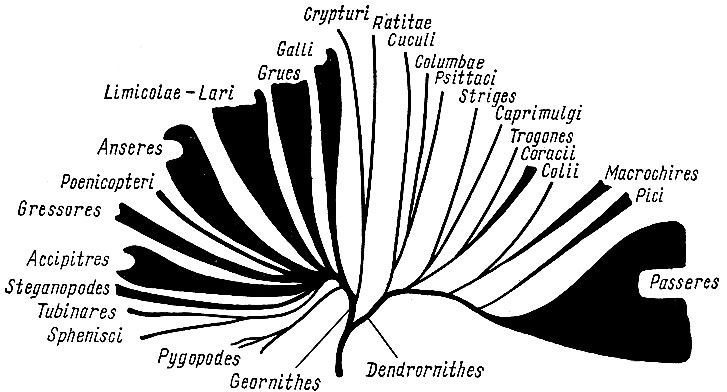
Рис. 15. Схема филогенетических отношений в классе птиц (по Берндту и майзе, 1960)
В последние годы попытку графического изображения филогенетических взаимоотношений между отдельными группами птиц предприняли немецкие орнитологи Берндт и Майзе в капитальной трехтомной сводке "Естественная история птиц" (Bernd R.,Meise W. Naturgeschichte derVogel, Kosmos, Stuttgart, 1960 (1962), v. 2, стр 668 - 673). Подкласс древних ящерохвостых птиц Palaeornithes дал начало подклассу новых веерохвостых птиц Neornithes, который включает 24 отряда, подразделяемых авторами обычно сразу на семейства; таксономические категории ранга подотряда и надсемейства используются, как правило, только в отряде воробьиных. Взаимоотношения между отрядами птиц, по Берндту и Майзе, выглядят следующим образом:
От древних птиц эволюция шла по двум направлениям (ветвям): ветви (надотряд) наземных и водных птиц Geornithes и ветви (надотряд) древесных птиц Dendrornithes. От самого начала ветви наземных и водных птиц (рис. 15) общим стволом отделились, позже разделившись, тинаму Crypturi и бескилевые птицы Ratitae, включая всех ныне живущих страусоподобных, в том числе и киви, и ископаемых моа и эпиорнисов. Далее обособляются куриные птицы Galli (в полном объеме отряда Уэтмора) и на этом же уровне, но самостоятельной ветвью - гагарообразные Pygopodes (включающие ископаемых меловых зубастых птиц гесперорнисов, гагар и поганок). Все остальные отряды этой ветви (надотряда) имеют общее происхождение - представляют собой как бы мутовку стволов, отходящих от одного участка. Относительно тесно связаны друг с другом и несколько противопоставляются другим группам отряды журавлеобразных (пастушки, мада- гаскарские пастушки, солнечные цапли, лапчатоноги, кагу, настоящие журавли, арамы, трубачи, дрофы, кариамы, трехперстки; из ископаемых - диатримы и форораки Phororhacidae) и болотно- и чайкообразных птиц Limicolae - Lari (яканы, различные группы собственно куликов, чайки чистики и - из ископаемых - зубастые меловые птицы - ихтиорнисы). Далее от общего основания мутовки (т. е. имея общих предков) последовательно обособляются отряды гусеобразных Anseres (паламедеи и собственно гусеобразные), фламинго Phoenicopteri, голенастые Gressores, дневные хищники Accipitres и веслоногие Steganopodes. От форм, близких к предкам веслоногих, отделяется ветвь, которая затем разделяется на отряды трубконосых Tubinares и пингвинов Sphenisci.
Развитие второй ветви (надотряда) древесных птиц Dendrornithes начинается от каких-то предковых форм, видимо, более древних, чем предки первой ветви (первого надотряда). Сначала общим стволом обособляются 3 отряда: кукушкообразных Cuculi, голубей Columbae (голубки, рябки и дронты) и попугаев Psittaci. Следующая обособившаяся ветвь разделяется на 2 ответвления, каждое из которых образует по 2 отряда. Одно из них дает начало совам Striges и козодоям Caprimulgi, другое - трогонам Trogones и ракшеобразным Coracii. Дальше от общего ствола также общей ветвью обособляются отряды птиц-мышей Colii и стрижеобразных Macrochires (стрижи и колибри), самостоятельной ветвью отходит отряд дятлов Pici (в объеме Уэтмора), а основной ствол ветвится на многочисленные подразделения отряда воробьиных Passeres. Помимо этого, авторы дают гипотетические схемы, в которых пытаются представить филогенетические взаимоотношения в пределах нескольких отдельных отрядов (дневных хищников, веслоногих, гагарообразных и воробьиных).
Филогенетические схемы Берндта и Майзе ясны, наглядны и легко запоминаются. Однако они практически совсем не аргументированы, не обоснованы материалами и, видимо, в значительно меньшей степени отражают действительные родственные (филогенетические) отношения между отдельными группами птиц, чем соответствующие схемы Фюрбрингера. Теоретически маловероятно, что уже на начальных стадиях эволюции класса четко определилась последующая биотопическая специализация птиц, что выражается в изложенной схеме деления подкласса "новых птиц" только на 2 обособленных надотряда и в почти линейном, последовательном обособлении отрядов. Оценивая по современным формам присущий птицам характер эволюции, выражающийся в широкой адаптивной радиации в пределах отдельных групп (о чем кратко упоминалось выше), кажется более логичным предполагать, что подобная же широкая адаптивная радиация была характерна и для начальных фаз эволюции класса. Если это так, то можно говорить, что филогенетические схемы Фюрбрингера хотя бы внешне действительно отражают преобладание такого типа эволюции.
Сейчас еще рано говорить о вполне сложившейся системе класса. Однако системы Фюрбрингера, Гадова и Уэтмора, видимо, могут рассматриваться как хорошая основа для построения естественной системы. Их материалы дают и самое общее представление о филогенетических взаимоотношениях большинства групп. Для улучшения и уточнения системы класса птиц необходимы дальнейшие разнообразные исследования на всех таксономических уровнях (от подвидовых группировок до отрядов и надотрядов): морфологические, экологические, этологические, генетические. Возможно, в будущем ценные материалы даст применение биохимических и физиологических показателей; использование современных методик пока существенного вклада в таксономию птиц не внесло. Хочется надеяться, что увеличится и количество палеонтологических материалов, которые позволят более обоснованно обсуждать эволюцию птиц в целом и отдельных групп (отрядов, семейств).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'