Происхождение новых видов
Объяснение происхождения видов - одна из основных проблем систематики. Состояние этой проблемы подробно изложено Э. Майром (1968).
С современной точки зрения, эволюционные изменения не обязательно приводят к увеличению числа видов. Например, изолированная популяция или группа популяций с течением времени может изменяться так, что вид "а" превратится постепенно в вид "d", пройдя эволюционные стадии видов "b" и "с" (рис. 6, А). Такие эволюционные преобразования, видимо, свойственны видам, имеющим ограниченные ареалы, что облегчает широкое перекрестное скрещивание выживших в результате отбора особей. Поэтому непрерывное генетическое изменение популяций, слагающих данный вид, может не сопровождаться возникновением репродуктивной изоляции между ними и не приводит к дивергенции - разделению первоначального вида на два или несколько новых.
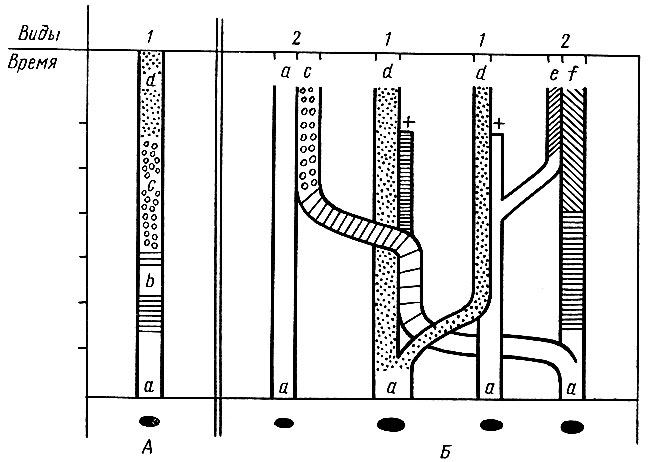
Рис. 6. Разные случаи видообразования по Майру (1968): А - изолированный остров, на котором вид 'а' превращается со временем в вид 'd', последовательно проходя стадии видов 'b' и 'с'; Б - архипелаг из четырех островов, на которых вид 'а' путем географического видообразования и колонизации образует 5 новых видов (и еще один - '6', вымирает), а на одном из островов сохраняется в неизмененном состоянии
Как происходит увеличение числа видов? У высших животных наиболее вероятным сейчас представляется постепенное видообразование, являющееся результатом постепенной дивергенции (расхождения) популяций. Видимо, основной путь возникновения новых видов - это географическое видообразование. Популяция, попавшая в изолированные условия и не подвергающаяся поэтому притоку генов из соседних популяций, под воздействием определенных внешних факторов проходит основной этап видообразования: перестройку генного комплекса всей популяции. Эта перестройка постепенно приводит к выработке механизмов, репродуктивно изолирующих данную популяцию от других близких популяций. При этом иногда изолирующие механизмы могут вырабатываться раньше и скорее, чем приобретаются морфологические различия. Когда изоляция прекращается из-за устранения преграды (отступание ледника, облесение луговых участков, соединение острова с другим островом и т. п.) или впоследствии происходит расселение с колонизацией новых территорий, эти новые виды могут вновь проникнуть на территории, занятые предковым видом, но выработавшиеся в период обособленного существования механизмы репродуктивной изоляции приведут к тому, что они встретятся как совершенно самостоятельные виды с симпатрическим распространением (рис. 6, Б).
Вероятно, иногда можно говорить об усилении изолирующих механизмов при образовании области совместного обитания двух близких видов, возникшей при расселении обоих видов или одного из них. Сейчас общеизвестна широкая географическая изменчивость песни у многих видов птиц. В ряде случаев различия в песне у симпатрических (встречающихся в одном районе) популяций двух близких и сходных по внешнему облику видов значительно более резкие, чем у популяций этих же видов, географически не сталкивающихся. На всех территориях, где совместно встречаются пеночка весничка Phylloscopus trochilus (L.) и пеночка теньковка Phylloscopus collybitus (Vieill.), их песни резко отличаются. На Пиренейском полуострове пеночки веснички нет; песня живущего здесь испанского подвида пеночки теньковки Ph. collybitus ibericus Ticehurst похожа на песню веснички и явно отличается от песни других подвидов теньковки (Тайсхерст, 1938, из Майра, 1947).
Наконец, два (или даже больше) вида могут возникнуть из широко распространенного политипического вида, когда его конечные популяции достигли или почти достигли репродуктивной изоляции, а соединявшие их промежуточные популяции вымерли по каким-либо причинам: изменению среды обитания (вторжение ледника или морского залива, появление или исчезновение лесов на больших территориях из-за изменения климата и т. п.), может быть, из-за захватившей обширную территорию интенсивной эпизоотии и т. д.
В хорошо изолированных районах (горные хребты, острова и т. п.) иногда встречаются два или более близких вида широко распространенной группы (обычно рода), которая в любом другом районе представлена лишь одним видом. Такие случаи часто рассматривают как доказательство симпатрического видообразования, т. е. дивергенцию одного вида на два в пределах одного и того же района. Ошибочность такой точки зрения была показана Э. Майром (1968) и другими исследователями; сейчас такие факты обычно объясняют иначе. Изолированный район был колонизирован группой особей, которая, развиваясь в изоляции, дивергировала от исходной родительской популяции и выработала изолирующие механизмы и экологическую обособленность к тому времени, когда в этот район вновь проникла следующая группа колонистов той же родительской формы. Такие двукратные и даже многократные волны нашествий выявляются при анализе состава фауны птиц ряда островов и даже континентов, например Австралии. Многократные переселения с острова на остров в пределах одного архипелага в некоторых случаях приводят к заметному увеличению числа видов и явному обогащению фауны. Сейчас не вызывает сомнения, что именно таким путем от 1 - 2 исходных предковых видов образовалось семейство (по представлениям части систематиков - подсемейство семейства Fringillidae) дарвиновых вьюрков Geospizidae на Галапагосских островах (14 видов, 21 подвид) и экологически и морфологически чрезвычайно разнообразное семейство гавайских цветочниц Drepanididae (45 подвидов, группируемых в 22 хороших вида).
Возможность симпатрического видообразования, т. е. возможность генетической дивергенции экологически специализированных популяций, населяющих одну и ту же территорию, без географической изоляции теоретически, видимо, допустима. Однако безупречных доказательств такого рода пока нет. Сейчас многими работами хорошо показано, что этому прежде всего препятствует расселение особей соседних, хотя экологически и разобщенных популяций, приводящее к генетической нивелировке подобных популяций и препятствующее видообразованию. Все факты, трактовавшиеся как случаи симпатрического видообразования, пока гораздо убедительнее и яснее объясняются с позиций географического видообразования (во всяком случае, применительно к птицам).
Образование нового вида путем слияния (гибридизации) двух ранее самостоятельных видов, видимо, в некоторых случаях возможно, но, вероятно, происходит крайне редко. Возможно, начало такого слияния видов представляет широкая гибридизация между двумя видами красноглазых тауи (Pipilo erythrophthalmus и P. ocai) во многих районах Мексики, вызванная, как предполагает ряд исследователей, резким изменением экологических условий в результате расширения сельскохозяйственных угодий за последние 300 - 500 лет и сильного сокращения площади естественных местообитаний этих видов. Остальные описанные случаи "слияния" видов в действительности представляют собой полосы гибридизации подвидов, без должных обоснований рассматриваемых как самостоятельные виды (Майр, 1968).
В очень редких случаях изоляционные механизмы между симпатри- ческими видами могут нарушаться в некоторых районах их совместного обитания.
Два близких вида воробьев - домовый Passer domesticus (L.) и испанский, или черногрудый P. hispanioletisis (Temm.), - встречаются вместе на обширной территории в Южной Европе и Западной Азии (рис. 7), не скрещиваясь друг с другом. На большей части ареала домовые воробьи живут преимущественно в населенных пунктах, а черногрудые заселяют рощи и кустарниковые заросли в поймах рек и по берегам озер. В нескольких районах - Южной Италии, на островах Сицилия, Корсика, Крит и в Тунисе - оба вида живут в культурном ландшафте и между ними идет гибридизация. В гибридных популяциях фенотипически обычно представлены оба родительских вида и все возможные комбинации родительских признаков, что свидетельствует о неограниченном случайном спаривании. Однако в некоторых локальных районах (Южная Италия, оазисы на юге Туниса) закрепился один из промежуточных гибридных типов, а родительские типы исчезли (по Майзе, 1936, из Майра, 1968). Во влажных тропических лесах Африки обычны два вида райских мухоловок Terpsiphone rufiventer и Т. rufocinerea, а во вторичных и саванных лесах - третий вид Т. viridis. Встречаясь в одних и тех районах, каждый из видов придерживается своего биотопа, и гибридизация между ними не отмечена. Однако в северо-западной Анголе, в Западной Африке и Уганде, где в последнее время влажные леса были частично или полностью сведены, Т. viridis скрещивается с обеими видами, особенно с Т. rufiventer, местами образуя устойчивые гибридные популяции (Чэпен, 1948, из Майра, 1968).
Подобные случаи нарушения видовых изолирующих механизмов, видимо, определяются резкими изменениями экологических условий среды обитания, прошедшими в относительно короткое время преимущественно в результате хозяйственной деятельности человека. Резкие изменения среды обитания, видимо, нарушили видовые стереотипы поведения и тем самым способствовали ослаблению и нарушению механизмов видовой репродуктивной изоляции.
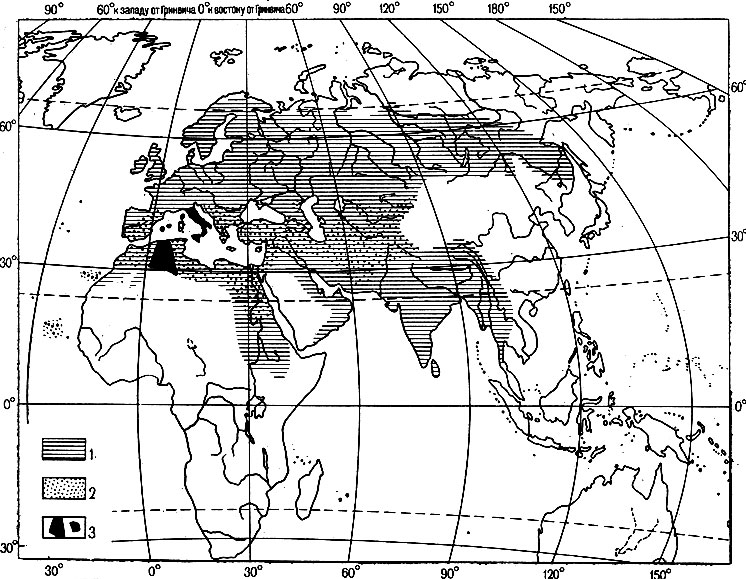
Рис. 7. Распространение воробьев: домового Passer domesticus (L.) и черногрудого P. hispaniolensis (Temm.) Майзе, 1936: 1 - арэал домового воробья, 2 - черногрудого, 3 - района гибридизации этих видов
Случайная межвидовая гибридизация отмечена у довольно большого числа видов птиц. Она установлена у многих видов уток (внутриродовые и даже несколько случаев межродовой гибридизации), куриных (между глухарем и тетеревом, тетеревом и белой куропаткой, тетеревом и рябчиком, между американскими луговыми тетеревами Tympanuchus cupido и Т. phasianellus, между некоторыми видами фазанов), дятлов (в частности, для видов рода Dendrocopos), ряда видов колибри, многих видов воробьиных (между чижом и щеглом, домовым и полевым воробьем, между рядом видов райских птиц, манакинов и др.) и некоторых других видов.
Такие случайные гибриды чаще встречаются у видов, которые не образуют пар или образуют их только на короткий период спаривания: кратковременность контактов самца и самки увеличивает возможность "ошибки" в выборе партнера (утки, многие куриные, колибри и др.). Для многих родов райских птиц Paradiseidae характерны групповые тока самцов, на которых самки появляются на короткое время; строит гнездо и насиживает только самка. У большинства этих родов описаны межвидовые гибриды. У довольно многочисленных по числу особей видов рода Manucodia гибридов не обнаружено: у этого рода образуются пары, и самцы участвуют в насиживании и выкармливании птенцов. Случайная гибридизация чаще отмечается на периферии ареала одного из гибридизирующих видов, где популяция обычно сильно изрежена, и особи испытывают большие трудности в отыскании партнера своего вида. Видимо, этой же причиной и, одновременно, резким нарушением видового стереотипа поведения объясняются относительно частые случаи случайной межвидовой гибридизации в условиях неволи даже между видами, гибриды которых в природных условиях не встречены (некоторые виды фазанов, воробьиных и др.). Однако в целом случайные гибриды в природных условиях встречаются крайне редко. Например, за 50 лет из Новой Гвинеи вывезено более 100 тыс. шкурок райских птиц, среди которых выявлено лишь 30 - 50 гибридов (Майр, 1968).
Видимо, такие гибриды очень интенсивно отметаются отбором, и, вероятно, не играют никакой роли в эволюции видов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'