Кольцевые перекрывания ареалов
Примерно у десятка видов птиц с широкими ареалами и отчетливой политипичностью обнаружено перекрывание ареалов конечных звеньев (форм), что теперь часто называют кольцевым перекрытием ареалов: цепь интерградирующих подвидов образует кольцо или петлю, в которой концевые звенья (подвиды) перекрываются, населяя одну и ту же местность, но не скрещиваются в зоне перекрывания. Эти случаи можно трактовать как неполное или как незавершенное видообразование.
Обычно в качестве примера приводят большую синицу Parus major, распространенную по значительной части Евразии и образующую до 30 подвидов. Северные и западные районы ареала вида занимают зеленоспинные и желтобрюхие подвиды группы mаjor (рис. 4). Южную Азию населяют сероспинные и белобрюхие подвиды группы cinereus, а восток Азии - зеленоспинные и белобрюхие подвиды группы minor. Все эти три группы подвидов гибридизируют в зонах контактов, более или менее постепенно переходя одна в другую. Раньше полагали, что кольцевое перекрывание ареалов группы major и группы minor в бассейне Амура не сопровождается гибридизацией, поэтому последнюю группу подвидов иногда выделяли в самостоятельный вид Parus minor. Учитывая, что гибридизацию в бассейне Амура можно, вероятно, считать доказанной, видимо, правильнее относить все эти географические формы к одному большому политипическому виду Parus major (некоторые систематики дробят его на 3 - 4 вида).
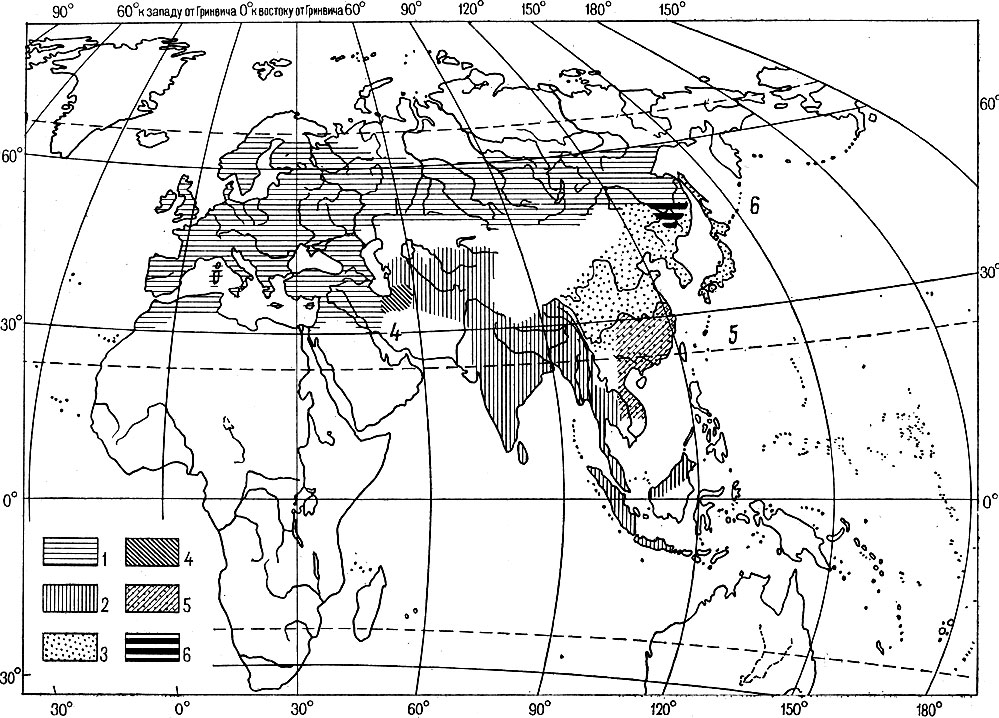
Рис. 4. Неполное видообразование у большой синицы Parus major (из Майр, 1968, по Делакуру и Вори, 1950; с изменениями): 1 - подвиды группы major (зеленая спина, желтое брюшко, из Западной и Северной Евразии), 2 - группы driereus (серая спина, белое брюшко, из Южной Азии), 3 - группы minor (зеленая спина, белое брюшко, из Восточной Азии); в зонах контактов образуются гибридные популяции: 4 - в Иране, 5 - в Южном Китае, 6 - в бассейне Амура. Есть предположение, что ареалы групп major и minor в бассейне Амура перекрываются без гибридизации
Бесспорнее другой пример кольцевого перекрывания ареалов: встречающиеся вместе на побережьях Балтийского и Белого морей и не образующие гибридов клуша Larus fuscus и серебристая чайка L. argentatus могут рассматриваться как перекрывающиеся концевые звенья цепи подвидов одного вида, широко распространенного в умеренных и северных широтах Евразии и Северной Америки. По представлениям изучавших этот случай Е. Штреземанна и Н. В. Тимофеева-Ресовского (1947, 1959), можно предполагать, что в доледниковое время серебристые чайки занимали обширный ареал. В плейстоцене широко распространившиеся ледники разделили ареал этого вида на несколько изолятов. Весьма вероятно, что центром формирования современных форм была не подвергавшаяся оледенению Берингия (стык ареалов форм vegae и smithsonianus). Отсюда в межледниковые и послеледниковую эпохи чайки расселялись сложным путем (рис. 5): на восток по Северной Америке (thayeri - leucopterus; географически сейчас обособленная califortiicus; argenteus - argentatus; последняя форма, видимо, относительно недавно проникла из Северной Америки в Северную Европу); на запад по северным побережьям Евразии (birulae - taimyrensis - antelius), через Западную Сибирь на юг (cachinnans - ponticus - armenicus) и оттуда на северо-запад (omissus) и через Средиземноморье в Атлантику (ponticus - michahellesi - atlanticus - britanicus - intermedius - fuscus). В результате в Беломоро-Балтийском районе сейчас встречаются четыре формы: fuscus, antelius (heuglini), omissus, argentatus (рис. 5), из которых первая (fuscus) морфологически и репродуктивно очень четко изолирована от остальных, а другие три морфологически отграничены довольно слабо, хотя гибриды между ними, видимо, довольно редки, и, кажется, намечается экологическая изоляция этих форм.
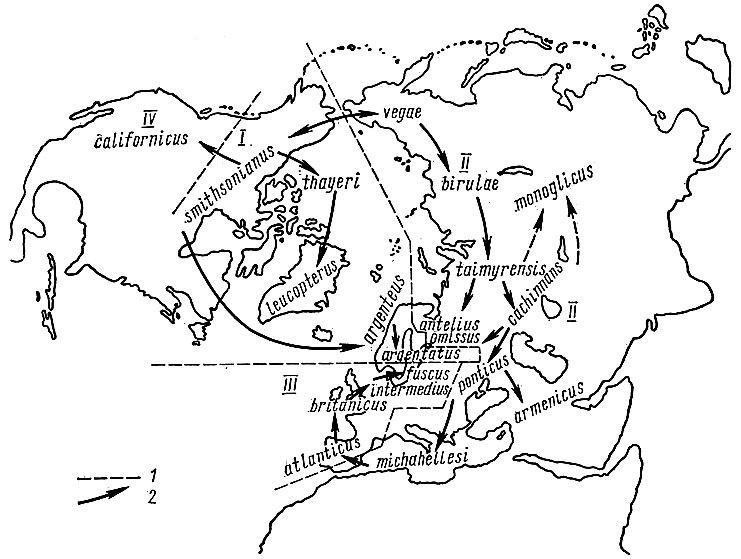
Рис. 5. Схема филогенетических отношений между географическими формами группы серебристая чайка - клуша (по Тимофееву - Ресовскому и Штреземанну, 1959). В Беломорско-Балтийском районе встречаются 4 формы: argentatus, antelius (heuglini), omissus, fuscus; из них наиболее обособлена клуша fuscus: I - границы между группами форм, иногда выделяемых в самостоятельные виды (I - серебристая чайка, II - чайка хохотунья, III - клуша, IV - калифорнийская чайка), 2 - предполагаемые пути возникновения и расселения отдельных форм
Поэтому при несомненной генетической близости всех упомянутых форм из-за перекрывания конечных звеньев и их репродуктивной изоляции в районе Балтика - Белое море сейчас обычно выделяют 2 вида: клушу L. fuscus (относя к нему формы fuscus, intermedins, britanicus, иногда atlanticus) и серебристую чайку L. argentatus со всеми остальными формами. Штреземанн и Тимофеев-Ресовский предлагают объединить эти родственные формы (подвиды) в 4 самостоятельных вида: клушу L. fuscus, серебристую чайку L. argentatus, чайку хохотунью L. cachinnans и калифорнийскую чайку L. californicus (распределение подвидов между этими видами показано на рис. 5). Некоторые авторы доводят число выделяемых видов до 5 и даже 6.
Этот пример хорошо показывает один из возможных путей видообразования, когда образовавшиеся виды обособились еще не полностью. Подобный тщательный систематический анализ ряда других крупных политипических видов несомненно углубит представления о путях и формах эволюционного процесса.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'