Перелет как физиологическая проблема
Как было установлено, проблема полета птиц на большой высоте требует более близкого рассмотрения. Проведенные ранее опыты относительно способности некоторых видов мелких птиц лететь на большой высоте кажутся бесполезными. Их результаты весьма незначительны, так как изучение реакции организма на уменьшение давления воздуха и понижение температуры возможно только в том случае, если животное находится в состоянии покоя, а это исключает сравнение с естественными условиями.
Напротив, при длительных пролетах птиц мы должны определять физиологические основы усиленной мышечной работы и ее источники. Это относится главным образом к таким видам, которые совершают тысячекилометровые перелеты над морем или в отношении которых удалось доказать, что они летят непрерывно в течение 12-15 час. и даже дольше. При 35-часовом пролете от Алеутских до Гавайских островов американская золотистая ржанка должна при двух ударах крыльев в секунду поднимать попускать их 252000 раз, совершая при этом огромную мышечную работу. Пролетный путь уже упоминавшейся полярной крачки, очевидно, самый продолжительный. Она следует из наиболее северной части своей гнездовой области, расположенной всего в 7° от Северного полюса, через Атлантический океан к берегам Европы и Африки и дальше к месту зимовок в Антарктике. Эти 17000 км птица должна покрыть в течение приблизительно 10 недель; следовательно, она должна ежедневно пролетать 225 км. Другие примеры продолжительных перелетов от 10000 до 15000 км у куликов также уже упоминались.
Когда были получены точные сведения о длительных перелетах, физиологи попытались согласовать их с данными об изменении обмена веществ. Однако до сих пор исчерпывающих результатов еще нет, и многие вопросы остались открытыми. Некоторые ученые считали, что непрерывный полет на тысячи километров и продолжительностью более 15-20 час. физиологически невозможен. Другие, напротив, пытались доказать, что необходимая для пролета энергия лишь незначительно превышает энергию, затрачиваемую птицей при ежедневных полетах в период размножения. Никольсон приводит несколько примеров: лазоревка, которая прилетает к гнезду только 35 раз в час, отлетая в поисках пищи на 100 м, покрывает 100 км в день. Ласточки или стрижи, весь день летающие со скоростью более 60-80 км/час, несомненно покрывают около 1000 км. По мнению этого автора, энергия, необходимая для миграции, соответствует энергии, затрачиваемой в повседневной жизни, но только направлена на определенную цель.
К некоторым видам особенно сильных летунов эти рассуждения, возможно, вполне подходят, но общего значения они, очевидно, иметь не могут, ибо физиологическая перестройка организма многих и даже большинства птиц незадолго до перелета и во время миграции указывает на то, что организм рассчитывает на совершенно исключительную нагрузку, в связи с чем изменяется его обмен веществ. Наиболее наглядно это проявляется в сильном накоплении жира, начинающемся вскоре после летней линьки и заканчивающемся за несколько дней или недель до начала осеннего перелета. Подобное явление наблюдается и к началу весеннего перелета. В общем птицы становятся тем более упитанными, чем длительнее предстоящая миграция и чем напряженнее она будет проходить. (В отношении крупных птиц данных еще нет.)
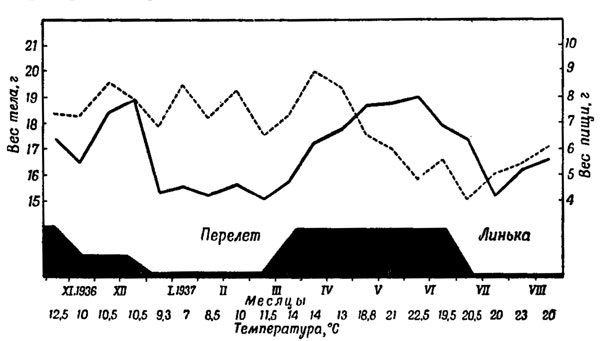
Рис. 35. Зависимость между весом тела, количеством пищи и 'перелетным беспокойством' у зарянки Erithacus rubecuia L. (Из Меркеля, 1940.) — вес тела; --- количество пищи
Это быстрое накопление жира до и в период перелета происходит за счет усиленного приема пищи. Однако одного этого было бы недостаточно - питательные вещества так же быстро расходовались бы. Физиологическая перестройка организма в сторону замедления обмена энергетических ресурсов и образования запасов, прежде всего - накопления жира, создает необходимые источники энергии. Если запасы жира после длительного полета израсходованы, птицы не в состоянии лететь дальше. Они остаются на богатых кормом местах до тех пор (иногда всего несколько дней), пока энергетические запасы вновь не будут пополнены.
Известно, что домашнего гуся особенно легко откормить осенью. Он является потомком совершающего дальние перелеты дикого серого гуся, у которого в периоды перелетов наблюдается склонность к накоплению жира. Став домашней птицей, гусь сохранил эту бесполезную для него физиологическую особенность перелетной птицы.
Еще несколько слов о зависимости между перелетом и сменой оперения у птиц. Оба процесса одинаково глубоко влияют на жизнь птицы, оба они в известной мере зависят друг от друга - один следует за другим,- о чем мы подробнее скажем ниже. Линька требует большой затраты энергии и поглощает все ресурсы организма. Это делает понятным, почему большинство птиц, особенно мелкие виды, не приступает к миграции во время линьки. Более крупные птицы в этом отношении менее чувствительны. Певчие птицы меняют маховые и рулевые перья до и после перелета. У них, помимо физиологических причин, решающее значение имеют летно-механические факторы (изъяны в несущих поверхностях). Рано отлетающие виды линяют обычно только на зимовках, поздно отлетающие - в гнездовой области. Часто мелкие перья сменяются на родине, крупные - только на зимовках (например, у городской ласточки, камышовок, пересмешек и др.). У крупных птиц эта зависимость между сменой оперения и перелетом в общем выражена менее четко, т. е. линька проходит медленнее и с большими промежутками.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'