Обмен веществ и энергии у перелетных птиц
Из сказанного выше можно представить такую картину. Сигнал из окружающей природы возбуждает один из центров гипоталамуса. Посылаемые им стимулы идут в двух направлениях: в расположенный рядом пищевой центр и в гипофиз. Пищевой центр резко увеличивает аппетит птицы, а гипофиз начинает перестраивать работу желез внутренней секреции. У оседлых птиц пищевой центр не получает такого приказа: естественный отбор не включил соответствующей информации в программу работы гипоталамуса.
Сильно возросший аппетит делает свое дело: перелетная птица начинает ежедневно съедать больше, чем тратит. Организм перегружается питательными веществами, которые отправляются на хранение в энергетические депо. Но птица располагает только жировыми депо, так как хранилища углеводов имеют очень ограниченную вместимость. Следовательно, все избыточные вещества начинают переводиться в жир. Превращение различных питательных веществ в жир контролирует промежуточный обмен, особенно интенсивный в печени. Постепенно жир начинает вытеснять гликоген отовсюду, где это возможно, - из печени, мышц. Промежуточный обмен веществ приобретает все более "жировой" характер. Углеводный обмен оттесняется на второй план. Такая ситуация очень затрудняет работу желез внутренней секреции. Одна из главных функций - управлять обменом веществ, удерживать его в определенных рамках, поддерживать постоянство внутренней среды. Но при перегрузке организма жирами и недостатке углеводов деятельность желез внутренней секреции затруднена. Обычно тонко уравновешенные антагонистические тенденции разных гормонов выходят из-под контроля и временно пересиливают то одни, то другие. Жировой и углеводный обмен тоже антагонистичны друг другу, но в нормальной обстановке гормоны "смиряют" противоречия, а теперь они проявляются в полной мере.
Одним из путей нормализации обмена веществ со стороны желез может быть усиление траты жира. Везде, где это возможно, ткани переходят на расходование жиров вместо углеводов. Увеличение расхода жира приводит к усилению двигательной активности птиц.
Но в результате этих изменений возникает еще большая неустойчивость внутренней среды. Возьмем для примера содержание сахара в крови. Обычно оно поддерживается на определенном, довольно постоянном уровне. А вовремя миграции стабилизация уровня сахара очень затрудняется: в организме сокращаются резервы углеводов, из которых можно срочно получить сахар, так как количество его в крови снижается. Если количество сахара в крови вдруг повысится, положение не улучшится. Раньше излишки сахара срочно передавались тканям. А теперь тканям не нужен сахар. Естественно, что уровень сахара в крови мигрирующих птиц будет очень сильно колебаться. Если уровень сахара низкий ("голодная кровь", как говорят физиологи), животное испытывает сильный голод, что способствует гиперфагии. При высоком содержании сахара ("сытая кровь") пропадает аппетит, но зато сильно возбуждается нервная система. Животное не спит, его все раздражает. Сложившаяся ситуация влияет на нервные центры, где сосредоточены врожденные пусковые механизмы миграционного поведения. Чувствительность центров повышается, и минимальный толчок запускает инстинктивное поведение. Птица начинает миграционный полет. Направление полета и поведение птиц запрограммировано во врожденном механизме точно так же, как в других нервных центрах запрограммировано умение строить гнездо или кормить птенцов.
Картина изменений, происходящих в теле птицы во время миграции, несколько упрощена. На самом деле, конечно, нарушение соотношения между жировым и углеводным обменом для птицы не неожиданное стихийное бедствие. Борьба между ними не бесконтрольный хаос, а скорее хорошо разыгранный спектакль, где каждый поворот действия тщательно отработан и давно предусмотрен естественным отбором (рис. 15).
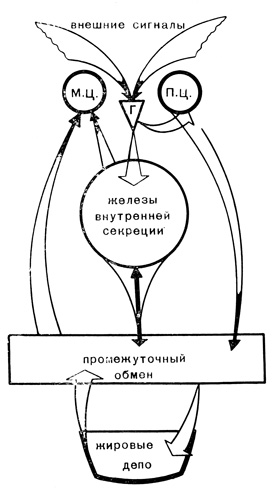
Рис 15. Схема возникновения перелетного состояния у птиц. Г - гипоталамус, П. Ц. - пищевой центр, М. Ц. - нервные центры, ответственные за миграционное поведение
Все это, конечно, тоже гипотеза, но она объединяет многие элементы гипотез, разбиравшихся выше. Можно ли подтвердить ее в опытах?
Осенью 1962 г. в Рыбачьем была начата обширная серия опытов по проверке этой гипотезы. Решено было взять за основу осеннее предмиграционное состояние - короткий период, в течение которого птицы переходят от линьки к миграции. Опыты проводились на зяблике - виде, наиболее удобном для работы в Рыбачьем. Основное внимание уделяли обмену веществ и поведению. Несколько сот молодых зябликов были пойманы и помещены в большие вольеры, построенные в тех местах, где птицы держатся в этот период года. Деревья в вольерах были оборудованы жердочками с контактами, которые позволяли регистрировать с помощью счетчиков изменения активности птиц. Такие же счетчики, помещенные на кормушках, давали представление о ритме питания. Чтобы разобраться в изменениях энергетического обмена в течение предмиграционного периода, часть птиц регулярно взвешивали и помещали в камеры, где измеряли количество поглощенного ими кислорода и выдыхаемого углекислого газа. Отношение углекислого газа к кислороду (респираторный коэффициент) отражает соотношение расхода углеводов и жиров в обмене веществ: если оно равно единице - организм расходует углеводы, если оно снизится до 0,7 - расходуются только жиры. Еще в 1958 г. Меркель отметил, что коэффициент выше единицы отражает очень важный процесс - интенсивное превращение углеводов в жир.
За соотношением жира и углеводов в обмене веществ следили и другим способом. Печень зябликов взвешивали, высушивали в термостатах до постоянного веса (чтобы удалить воду) и вновь взвешивали. Затем ее растирали в порошок и экстрагировали жир. У тех же птиц собирали в пробирки кровь и определяли содержание сахара. Чтобы следить за ходом усвоения питательных веществ, взвешивали содержимое желудка и кишечник. Железы внутренней секреции и гипоталамус фиксировались для последующего изучения их активности. После расчленения и взвешивания отдельных органов тушку высушивали и определяли в ней количество воды и жира.
Одновременно в природных условиях велись наблюдения за изменениями в кормежке и перелетным поведением свободноживущих птиц. Часть из них приходилось отстреливать, чтобы определить состав поедаемой пищи и сравнить жировой и углеводный обмен свободноживущих птиц с подопытными. Сотни птиц отлавливались ловушками. Перед выпуском их взвешивали, измеряли, описывали ход линьки и оценивали количество подкожного жира.
Сотрудники станции и приехавшие для выполнения этой обширной работы специалисты из других институтов проводили эксперименты круглосуточно, так как наиболее интересны изменения обмена веществ в течение суток. Чтобы лучше разобраться в происходящих изменениях, дополнительный материал собирался на всем миграционном пути зяблика - от Полярного круга до зимовок. Ценные сведения поступили из Лапландского заповедника, Кандалакши, Карелии, Ленинградской области, Пскова, Эстонии. Очень важное исследование провел профессор Шильдмахер на орнитологической станции Хиддензее на севере ГДР; поступили подробные данные о миграции зябликов из Южной Швеции, Бельгии, Франции и Швейцарии. Все эти коллективные усилия позволили подробно разобрать изменения в обмене веществ зябликов при наступлении миграционного состояния.
Что же происходит в организме зябликов перед началом миграции?
Опыты были начаты 20 августа - за декаду до конца линьки. В это время в природе зяблики держались небольшими стайками по 5-20 птиц и кормились. Кормежка начиналась с восходом солнца и была особенно активной около 10 часов утра (рис. 16). В полуденные часы они кормились не так активно и многие просто отдыхали, а ближе к вечеру вновь принимались за поиски пищи. В течение всего дня стаи не раз перелетали с места на место в поисках корма и воды. Подобным образом вели себя и зяблики в вольерах: вся их активность была направлена на кормежку, поэтому в те часы, когда счетчики регистрировали максимум прыжков между деревьями, счетчики на кормушках тоже отмечали высокую активность. Несмотря на обилие съедобных семян, птицы на воле предпочитали поедать насекомых. В одной и той же стае линяющие птицы наполняли свои желудки только насекомыми и зеленью; птицы, заканчивающие линьку, животную пищу дополняли семенами, а у птиц, окончивших линьку, в желудке были почти одни семена. Такая избирательность питания отражает различие в потребностях линяющих птиц и уже отлинявших: первым нужно много белков, солей и витаминов для обновления тканей, а вторые предпочитают богатые углеводами и жирами семена.
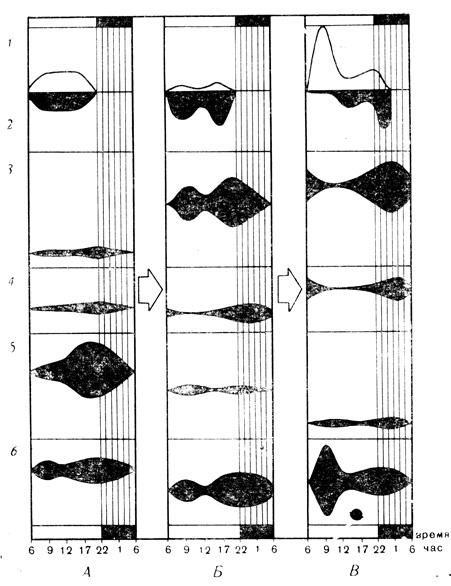
Рис. 16. Осенний предмиграционный период у зяблика. А - конец линьки, Б - предмиграционное накопление жира, В - начало миграции; 1 - активность зябликов, 2 - кормежка, 3 - уровень жировых резервов в теле, 4 - жир печени, 5 - резервы гликогена в печени, 6 - уровень сахара в крови
Съеденная и усвоенная пища переводилась в углеводы, которые накапливались в печени в течение дня, а ночью расходовались: вес ее увеличивался к ночи, а к утру уменьшался. Респираторный коэффициент подтверждал, что основным источником энергии в организме птиц были углеводы. Жира в теле зябликов и в печени было мало, и он не проявлял никакого антагонизма по отношению к углеводам: подобно им, он накапливался днем и расходовался ночью.
В конце августа - первых числах сентября поведение зябликов в природе постепенно изменилось: стаи увеличились до нескольких десятков особей. Они по-прежнему кормились весь день, но вечерняя кормежка становилась все более активной и продолжалась до самого захода солнца. Изменился и состав пищи: зяблики начали поедать в основном семена. И в природе, и в вольерах сильно снизилась подвижность птиц. В вольерах она была в семь раз ниже, чем в конце линьки! Кроме активности, связанной с поиском корма, в утренние часы постепенно появлялась независимая от кормежки активность: птицы без видимой цели перелетали из конца в конец вольеры, перекликались. Это было уже проявлением слабого перелетного беспокойства. К середине сентября утреннее беспокойство все более усиливалось, а кормежка в эти часы ослабевала. Еще в самом начале этого периода изменился характер углеводного обмена. Сначала углеводов в печени было столь же много, как и в конце линьки, но они не накапливались днем и почти не тратились ночью: количество их оставалось постоянным в течение суток. Но день ото дня их становилось в печени все меньше, и вес печени соответственно уменьшался. Количество воды в теле уменьшилось на один грамм. Жира сначала было не больше, чем в конце линьки, но количество его сильно изменялось в течение суток, ночью он активно расходовался, а днем накапливался в еще больших количествах. День ото дня, несмотря на усиленную трату, количество его в организме возрастало. Начался активный синтез жира. Респираторный коэффициент после кормежки достигал 1,4-1,7, что отражало перевод углеводов в жиры.
Источником начавшегося ожирения было переедание. Если энергия, усвоенная из пищи, во время линьки была равна затратам, то в начале предмиграционного периода она превышала их на 0,15 ккал в сутки, а в конце - на 4,1 ккал! К концу предмиграционного периода углеводный обмен был подавлен, а резервы углеводов почти полностью ликвидированы. Жировой обмен, напротив, возрос и стал господствующим. Жировые резервы достигли высокого уровня, свойственного миграции. Все измерявшиеся признаки приобрели один из двух противоположных суточных ритмов. Если раньше ритмы кормовой активности, подвижности, жировых резервов и сахара крови были одинаковы, то теперь они стали антагонистичны.
Утром 16 сентября к югу потянулись огромные стаи: началась миграция. К этому же времени утреннее беспокойство птиц в вольерах достигло апогея: оно в 12 раз превышало активность в предмиграционный период. Суточный ритм сахара крови не изменился, но уровень его стал очень неустойчивым: утром, после пробуждения, он стремительно повышался, достигая очень высокого уровня. В эти часы птицы не кормились ("сытая кровь"), но сильно беспокоились, а в природе - летели. Жировые резервы интенсивно расходовались, респираторный коэффициент был низок (сжигался жир). Так продолжалось до полудня. После полудня и в природе и в вольере миграционное поведение прекращалось, сахар крови стремительно снижался до очень низкого уровня ("голодная кровь") и начиналась активная кормежка. Респираторный коэффициент в это время был выше единицы: углеводы пищи переводились в жир. Жировые резервы начали возрастать.
О чем говорят результаты этих опытов? Они, несомненно, хорошо соответствуют гипотезе, для проверки которой они и были проделаны. Но для того чтобы полностью проверить ее, нужны исследования на многих видах, проведенные в высоких широтах и низких, весной и осенью. Мы должны убедиться, что перелетное состояние имеет место всякий раз, когда угнетается углеводный обмен и усиливается жировой, и наоборот, оно не возникает никогда, если между ними нет антагонизма.
Летом и осенью 1963 г. было исследовано еще три вида: садовые славки, зарянки и скворцы. Славки - ночные мигранты, у них перелетное и кормовое поведение не сталкиваются. Как в таком случае обстоит дело с жировым и углеводным обменом? Оказалось, что и у них в период миграции резко усиливается жировой обмен. Зарянки - тоже ночные мигранты, но у них, как и у скворцов, есть одна особенность - они часто мигрируют осенью без больших жировых запасов. Оказалось, что у этих двух видов птиц жировой обмен усиливается, а углеводный угнетается. Несмотря на небольшой уровень жировых резервов использование жира в течение суток при переходе к миграции резко возрастает; в печени гликоген замещается жиром.
В том же году был изучен обмен веществ еще у одного вида - чижей. Сравнили три состояния: характерный для этого вида промежуточный перелет в северном направлении (после размножения перед линькой), линьку и осеннюю миграцию. Промежуточный и осенний перелеты, в отличие от линьки, были связаны с соответствующими изменениями в жировом и углеводном обмене.
Различия между обменом веществ в период линьки и во время осенней миграции очень велики: для линьки характерен углеводный обмен, а для миграции - жировой. Но на севере птицы вынуждены совмещать линьку с началом отлета на юг. Так обстоит дело у пеночек-весничек - небольших насекомоядных птичек, гнездящихся вплоть до Полярного круга, а в конце лета улетающих на зимовку в Африку. В средних широтах, например в Рыбачьем, они линяют чрезвычайно интенсивно и миграция начинается после окончания линьки. Наступление миграционного состояния связано у них с усилением жирового обмена. А на севере, например в Карелии, где отлет начинается уже во время линьки, уровень жировых резервов в теле и в печени все равно возрастает и трата жира усиливается. Необходимый для линьки углеводный обмен остается на прежнем уровне, т. е. в этом случае наблюдается абсолютное усиление жирового и относительное ослабление углеводного обмена.
До сих пор изучались перелетные виды. Не менее интересны отношения жирового и углеводного обмена весной и осенью у оседлых видов. К сожалению, среди наших птиц почти нет удобных для наблюдения оседлых видов. Практически - одни воробьи. В 1963 г. изучалось осеннее состояние у домовых и полевых воробьев. Оказалось, что у них осенью жировой и углеводный обмен в равной степени активны и не противопоставлены друг другу. Но, к сожалению, воробьи сильно отличаются от всех наших птиц: они появились на севере сравнительно недавно (с человеком). Птицы эти тропического происхождения, сравнивать их с северными птицами трудно. Но и из этого положения нашелся выход: в Средней Азии летом обитает близкий подвид нашего домового воробья. Это перелетный воробей: к концу лета, перелиняв, птицы собираются в огромные стаи и летят через горы, в Индию, где зимуют.
Осенью 1964 г. в Таджикистане были исследованы перелетные воробьи. Оказалось, что невзирая на тропическое происхождение они перед отлетом начинают жиреть и жировой обмен у них угнетает углеводный - совсем как у других перелетных птиц. Значит, отсутствие антагонизма между жировым и углеводным обменом у оседлых воробьев не случайность и не результат их тропического происхождения. К этому времени в Индии ученые проверяли соотношение жирового и углеводного обмена у зимующих там розовых скворцов. У этих птиц подготовка к перелету на север тоже сопровождалась угнетением углеводного обмена и усилением жирового.
Итак, за три года обследовано довольно много видов, и всякий раз результаты соответствовали гипотезе. Но в природе около 8000 видов птиц, и среди них могут вполне оказаться такие, миграционное состояние которых не будет соответствовать гипотезе. Изучить все виды, конечно, никто не может. Нужно проверять гипотезу другим путем.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'