Дополнительная энергия перелета
Чтобы понять, откуда птицы берут дополнительную энергию, которую они запасают в жире, нужно выяснить, как расходуется энергия, заключенная в пище (так называемая большая энергия). Во-первых, не вся пища усваивается организмом, часть ее выбрасывается наружу непереваренной. У здоровой птицы, питающейся доброкачественной пищей, усваивается 70-80% большой энергии. Некоторая часть этой "усвоенной энергии" расходуется на пищеварение и усвоение пищи, а оставшаяся часть - на удовлетворение других нужд организма - "метаболизированная энергия". Но энергия, расходуемая организмом на жизнедеятельность - "катаболизированная энергия", не всегда равна количеству энергии, поступившей с пищей. Если она больше метаболизированной энергии, то недостаток покрывается за счет использования энергетических резервов, а если меньше, то избыток откладывается в форме резервных питательных веществ. В первом случае энергетический баланс положителен, а во втором - отрицателен. Конечно, накопление жира может идти только при положительном балансе, т. е. лишь в том случае, когда птица получает из пищи больше энергии, чем расходует. Итак, вопрос об источниках дополнительной энергии птиц в миграционный период сводится к выяснению источников, обеспечивающих положительный энергетический баланс.
Положительный баланс может быть обеспечен за счет сокращения трат организма, за счет увеличения количества поедаемой пищи и, наконец, в результате совмещения того и другого. Долгое время в науке преобладала первая точка зрения, которую наиболее четко сформулировал еще в 1932 г. немецкий ученый Грёббельс. Он предположил, что птицы во все сезоны года поедают примерно одинаковое количество пищи. Зимой полученная из корма энергия расходуется на поддержание температуры тела, летом - на размножение и смену перьев. Весной и осенью энергетические затраты птиц меньше и в организме образуется жир.
Несмотря на простоту эта гипотеза не может быть принята: исследования энергетического обмена птиц в разные сезоны года не подтвердили ее. Потребность птицы в корме сильно меняется в разные сезоны. Она проявляется сильнее всего летом, когда в природе много пищи. К кормному периоду года приурочены как раз те фазы годового цикла птиц, которые требуют особенно много энергии. Меньше всего птицы нуждаются в корме зимой, когда его мало, так как зимой обмен "разгружен" от всяких затрат, кроме самых необходимых. Более того, чем севернее зимует птица, тем сильнее приспособления к суровой зиме и тем ниже у нее обмен. Это свойство птиц имеет огромное значение для миграций: если бы расход энергии не снижался зимой, не имело бы смысла улетать на зимовки. Зимнее снижение обмена у перелетных птиц, вместе с продемонстрированным недавно встречным снижением обмена у аборигенов, - важнейшее приспособление к миграции. Кстати это ответ на вопрос; почему перелетные птицы не остаются на зимовках навсегда? Это невозможно, потому что весной повышение потребности в пище у гостей и аборигенов вызывало бы конкуренцию.
К изложенной выше гипотезе близка точка зрения, согласно которой миграционное отложение жира - результат хороших кормовых условий весной и осенью (питания жирными кормами) или продолжительного кормового дня. С этой точки зрения проявление глубочайших приспособлений птицы к окружающему миру - результат стечения благоприятных обстоятельств.
Строго научные опыты показали, что ни состав пищи, ни короткий кормовой день не могут прекратить отложение жира у перелетных птиц или вызвать его у оседлых птиц. В немиграционный период ни обильная пища, ни продолжительный кормовой день, ни прочие способствующие положительному балансу условия не вызывают обильного накопления жира. Уровень жировых резервов птицы соответствует ее потребностям в данное время, и только отсутствие пищи может нарушить это соответствие.
В 1956 г. изучение биоэнергетики весеннего предмиграционного периода проводилось на американской овсянке Гамбеля (рис. 11). Подопытные птицы были посажены в конце зимы в клетки, где в изобилии получали пищу, количество которой и калорийность все время учитывались. Раз в несколько дней клетку чистили и определяли калорийность остатков пищи и экскрементов. Разность между количеством калорий, поступавших с пищей и удаленных из нее с остатками, соответствовала энергии, усвоенной птицами.
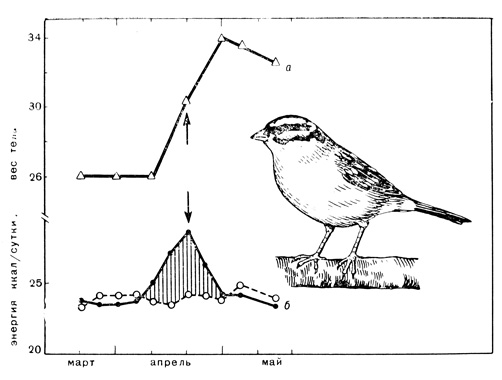
Рис. 11. Опыты с овсянкой Гамбеля; а - изменение веса овсянки перед весенним отлетом на родину; б - изменение энергии; сплошная линия-энергия, полученная с пищей (метаболизированная); прерывистая линия - энергия, израсходованная организмом (катаболизированная). Перелетное беспокойство (обозначено стрелкой) появилось после того, как птица отложила жир
В течение некоторого времени вес птиц и количество полученной ими энергии не менялись. Но как только наступила подготовка к весенней миграции, птицы стали поедать больше пищи, количество полученной из пищи энергии возросло на 30-40%, увеличился вес тела и началось отложение жира. Подобные данные получены уже более чем для 25 видов мелких птиц в период подготовки к весенней и осенней миграциям.
Итак, отложение жира основано на переедании (гиперфагии), которое возникает при любом - высоком и низком - уровне обмена. Но, конечно, если обмен ниже, отложение жира происходит быстрее. Поэтому у дальних мигрантов в период миграции часто обмен ниже, чем у оседлых птиц близких видов.
Накопление жира перед миграцией продолжается всего 10-15 дней, и за это время в теле птицы откладывается необходимый запас жира. Отложение жира у мелких птиц идет со скоростью 0,1-0,5 г в сутки.
Но вот, наконец, началась миграция, птицы полетели, и как ни велики резервы их энергии, они все же истощились и произошло это на пути к цели. Что дальше? Птица временно прекращает миграцию и начинает настойчиво кормиться. Гиперфагия вновь проявляется с полной силой, и жировые резервы постепенно восстанавливаются. Отощавшая птица при обилии корма в клетке может накапливать до 2-5 г жира за сутки! Скорость накопления жира у птиц в естественной обстановке до недавнего времени была неизвестна. Но в последние годы ученые собрали очень ценные данные о накоплении жира у птиц во время миграции. Выяснилось, что в отдельных случаях повторно пойманные окольцованные птицы накапливают за сутки более 1 г жира. У птицы размером с воробья за сутки накапливается около 0,5 г жира.
В 1963 г. изучили процесс накопления жира у рекордсмена беспосадочных полетов - американской древесной славки. Эта птица, весящая 12 г, живет на севере Америки, а зимует в Венесуэле. Осенью по дороге на зимовки 100 млн. птиц появляются на северо-востоке США, в штате Массачусетс, куда прилетают ночью, отощавшие после броска. Здесь древесные славки в течение трех недель кормятся, накапливая за это время 10 г жира. Затем в одну из ночей многомиллионная стая поднимается в воздух и улетает в сторону Атлантического океана, чтобы кратчайшим путем достигнуть Южной Америки. Столь быстрое восстановление резервов объясняется удивительной скоростью поедания и переваривания пищи птицами. Чижи или зяблики, например, даже поедая семена размером с маковое зернышко, набивают желудок за 15-20 минут (а ведь каждое зернышко нужно вынуть и очистить!). А через час их кишечник уже пуст: все переварено. Перееданию способствует и то, что тощие птицы неактивны, но много кормятся. Это очень целесообразно: попытка без резерва энергии перелететь море или пустыню равносильна гибели.
Не подошли ли мы к разгадке причин, вызывающих перелет? Сопоставим такие факты: оседлые птицы не откладывают жира весной и осенью, они и не мигрируют; перелетные же птицы накапливают жир. Перелетные птицы тем активнее, чем жирнее. Стоит им растратить жир, как их перелетная активность прекращается. Накопив жир, они вновь начинают перелет. Почему бы не предположить, что физиологическая основа миграционного состояния - большие жировые резервы? Именно так и полагал Грёббельс. Его гипотеза объясняла многие факты и существовала с 1932 г. без всяких изменений около 30 лет.
В последние годы было предложено еще несколько гипотез. Кенди и его коллеги в 1960 г. обнаружили, что у американской воробьиной овсянки перелетное беспокойство появляется до начала накопления жира и затем сразу наступает ожирение. Ученые предположили, что начавшееся перелетное поведение приводит к усилению траты жира и служит сигналом к его накоплению (рис. 12).
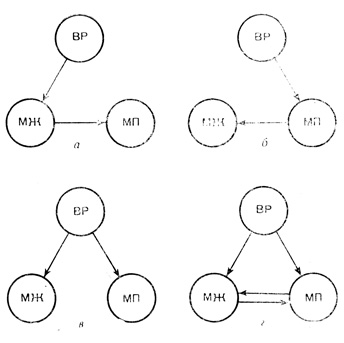
Рис. 12. Связь между отложением жира и миграционным поведением по разным гипотезам; а - гипотеза Грёббельса, б - Кенди, в - Кинга и Фарнера, г - новейшая. Миграционное отложение в теле жира - МЖ, миграционное поведение - МП, деятельность высших регуляторов - ВР
В 1963 г. Кинг и Фарнер пришли к выводу, что отложение жира в теле перелетных птиц и возникновение у них перелетного беспокойства - два независимых процесса, вызываемых общей причиной.
По мнению других ученых (В. Р. Дольник и сотрудники), ожирение и миграционное поведение - два параллельных процесса, влияющих друг на друга. Это предположение не отрицает взглядов других исследователей и рассматривает их как верное, но в каждом случае неполное описание действительных связей.
Какие же факты доказывают правильность каждой из нарисованных на рис. 12 связей? Начнем с доказательств того, что отложение жира и перелетное поведение включаются независимо. Во-первых, далеко не всегда обильное отложение жира приводит к миграции. Зимующие в высоких широтах птицы, как правило, зимой очень жирны. Этот жир нужен им как резерв энергии в течение длинной зимней ночи и на случай бескормицы из-за непогоды. И хотя жировые резервы зимой больше, чем во время миграции, зимующие птицы живут оседло и активность их очень мала. Во-вторых, некоторые виды птиц, например скворцы или зарянки, мигрируют с очень небольшими жировыми резервами. У всех видов птиц в пролетных стаях присутствуют отдельные тощие птицы. Также птицы тоже проявляют перелетное беспокойство. Это доказано на опытах. Экспериментаторы разделили подопытных птиц в конце зимы на две группы: одну кормили достаточно, а другую ограничили в пище настолько, что птицы не могли ожиреть. В результате с наступлением весны одни птицы отложили миграционный жир, а другие остались тощими. Но и у тех и у других появилось перелетное беспокойство, правда у тощих более слабое.
В другом опыте, проведенном зимой, зяблики были помещены в камеры, где поддерживался световой день различной продолжительности. При не очень длинном световом дне перелетное беспокойство появилось на много дней раньше, чем отложение жира. Итак, как ни тесно связаны перелетное поведение и миграционное ожирение, все же они регулируются независимо. Мы уже знаем, что у тощих птиц перелетное беспокойство выражено слабо, а у жирных - сильно. Что произойдет, если ввести в жировое депо тощих птиц жир? Жир не содержит ничего такого, что отравляло бы организм. Жиры разных животных имеют практически один и тот же качественный состав, они отличаются друг от друга только соотношением компонентов. Жир одного и того же вида птиц почти не отличается.
Ввести подопытным птицам жир очень просто: с помощью шприца прямо в подкожные жировые депо. В первых опытах вводился жир, экстрагированный из жирных зябликов. Зимой в автоматически регистрирующие активность и кормежку клетки были помещены зяблики и домовые воробьи. Две группы воробьев и две группы зябликов были в типичном для зимы состоянии, и одной из них ввели жир, другая служила контролем. Ни у оседлых воробьев, ни у перелетных зябликов введение жира не вызвало перелетного беспокойства: на птиц в зимнем состоянии жир не действовал. Тогда птицы были стимулированы к переходу в весеннее состояние длинным световым днем. Воробьи при, этом не ожирели, а зяблики в одной контрольной группе ожирели. Зябликам из двух других групп накопить жир не позволили, ограничив питание. Ни воробьи, ни тощие зяблики почти не беспокоились, в отличие от жирных зябликов, у которых появилось настоящее перелетное беспокойство. Тогда воробьям и одной группе зябликов ввели жир. На воробьев это опять не подействовало, а зябликов привело к настоящему перелетному беспокойству.
Итак, введение жира тощим перелетным птицам в период миграции усиливает миграционное поведение. На оседлых птиц жир не оказывает такого действия.
Но почему именно жир? Может быть, это какой-нибудь его спутник? Нет, теперь мы можем точно сказать, что это именно жир. В этом убеждает такой опыт: вместо птичьего жира зябликам вводили жир, экстрагированный из растительных масел, даже из обычного сливочного масла, - результат был одинаков. Значит, не состав жиров, не какая-то примесь, а комплекс жирных кислот, составляющих жиры, меняет перелетное поведение.
Роль жировых резервов очень велика, но само по себе накопление жира - еще не причина возникновения перелетного поведения птицы. Эту причину нужно искать в каких-то иных реакциях организма. Поиску этих загадочных стимулов, вызывающих дважды в год "потребность к перемене мест", мы и посвятим следующую главу.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'