Кинетизм черепа и типы строения костного неба
Костное небо птиц подвижно. Оно образовано небными отростками межчелюстных и верхнечелюстных костей, сошником, небными и крыловидными костями. Подвижные квадратные кости сочленяются с крыловидными костями, которые в свою очередь сочленяются с небными. Сочленение небных и крыловидных костей может скользить вдоль клювовидного отростка парасфеноида. Подвижность костного неба и истончение костей у основания верхней части надклювья в области "переносицы" (а иногда и образование здесь соединительнотканной перемычки - синдесмоза или даже настоящего сустава) обеспечивают кинетизм - способность надклювья смещаться вверх и вниз по отношению к мозговому отделу черепа. Для птиц характерна кинетичность двух типов (Юдин, 1965, 1970; Дзержинский, 1971): прокинетизм (гибкая зона лежит у основания надклювья) и ринхокинетизм (гибкая зона расположена на уровне носовых ям, верхняя челюсть обладает внутренней подвижностью). Кинетизм позволяет увеличить усилие на конце клюва, обеспечивает (вместе с дифферент циацией челюстных мышц и связок) разнообразие производимых клювом движений, облегчает заглатывание добычи.
Выделяют 4 типа костного неба, различающихся по расположению образующих его костей (рис. 11):
1. Дромеогнатический тип. Небные и крыловидные кости неподвижно соединяются друг с другом; хорошо развиты базиптеригоидные сочленения; передний конец сошника обычно сливается с небными отростками верхнечелюстных костей, задний конец сошника расщеплен и его расходящиеся концы соединяются с небными и крыловидными костями, отделяя их от клювовидного отростка. Различные вариации такого типа костного неба характерны для тинаму и страусоподобных бескилевых птиц.
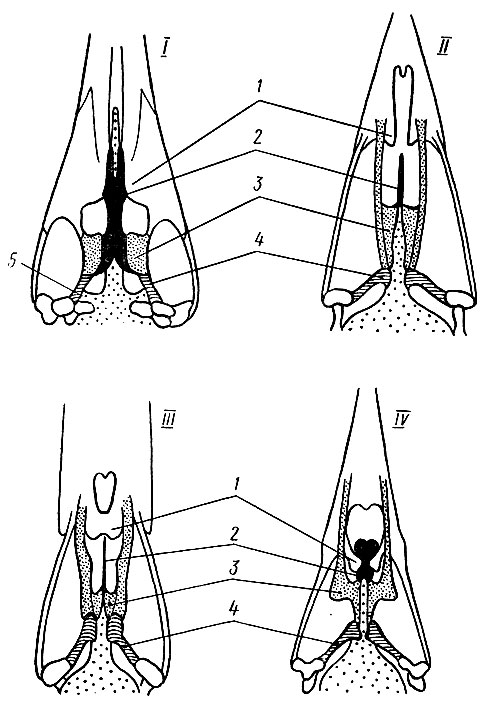
Рис. 11. Схема типов костного неба птиц. I - дромеогнатический (нанду); II - схизогнатический (курица); III - десмогнатический (утка); IV - эгитогнатический (ворона): 1- небные отростки верхнечелюстных костей, 2 - сошник, 3 - небные кости, 4 - крыловидные кости, 5 - базиптеригоидное сочленение
2. Схизогнатический тип. Небные отростки правой и левой верхнечелюстных костей не соединяются друг с другом; небные и крыловидные кости подвижно сочленяются друг с другом и примыкают к клювовидному отростку; сошник спереди заострен и свободен - не соединяется с небными отростками верхнечелюстных костей. Различные вариации схизогнатического типа неба характерны для пингвинов, гагар, трубконосых, куриных, большинства журавлеобразных, ржанкообразных, трогонов, большинства сов, козодоевых, колибри, настоящих дятлов.
3. Десмогнатический тип. Небные отростки правой и левой верхнечелюстных костей сливаются друг с другом; сошник либо совсем редуцируется, либо узкий, палочковидный, задним концом иногда соединяющийся с небными костями и клювовидным отростком, а спереди - со слившимися небными отростками верхнечелюстных костей. Вариации такого типа неба свойственны веслоногим, голенастым, гусеобразным, дневным хищникам, некоторым козодоям, ракшеобразным, якамарам, пуховкам и туканам из дятлообразных.
4. Эгитогнатический тип. Тонкие удлиненные небные отростки верхнечелюстных костей подходят под сошник (если смотреть на череп снизу, то они нависают над сошником), но не соприкасаются ни с сошником, ни друг с другом; сошник широкий со свободным широким, иногда раздвоенным передним концом; задний его конец глубоко расщеплен и охватывает клювовидный отросток. Такой тип неба встречается у трехперсток, стрижей, бородаток и медоуказчиков из дятлообразных, почти у всех воробьиных.
Второй - четвертый типы неба иногда объединяют в группу неогнат (новонебных), противопоставляя ее дромеогнатическому или палеогнатическому (древненебному) типу. Десмогнатический иэгитогнатический типы неба, видимо, возникли путем перестройки более древнего схизогнатического типа. Между всеми этими типами неба существуют более или менее выраженные переходы.
Базиптеригоидные сочленения. У страусоподобных птиц, тинаму, некоторых трубконосых, гусеобразных, сов и некоторых других птиц на боковых сторонах основной клиновидной кости расположены выросты - базиптеригоидные отростки; каждый из них образует скользящее сочленение с боковой поверхностью крыловидной кости соответствующей стороны. У пингвинов, чаек, чистиковых и некоторых других базиптеригоидные сочленения образуются в эмбриогенезе и полностью исчезают по мере роста птенцов.
У трубконосых, веслоногих, чаек и других птиц имеется стрептогнатизм: благодаря специальным устройствам ветви нижней челюсти (подклювья) могут выгибаться наружу, значительно увеличивая раскрытие ротового отверстия и обеспечивая заглатывание крупной добычи (Юдин, 1965, 1970).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'