Глава 17. Эксперименты по узнаванию яиц
Реакция птиц па яйца открывает широкие возможности изучения внешних раздражителен, которые воздействуют на птицу, так как с яйцами легко манипулировать; к тому же они в отличие от птенцов и взрослых птиц сами двигаться песпособны и их можно заменять точными моделями. Это позволяет изучить воздействие различных признаков яйца, таких, как, скажем, цвет или форма, сравнивая реакцию птиц на точные модели с их реакцией на модели, в которых то или иное свойство опущено или изменено. Во всяком случае, анализ системы раздражителей, определяющих поведение в период насиживания, - одна из первых задач, с которыми сталкивается наблюдатель, так как способность чаек находить свое гнездо среди множества других представляет собой одно из поразнтельиейших явлений в жизни колонии. Это особенно изумляет туриста-горожанина: сам он очень плохо ориентируется в дюнах, где нет улиц с табличками их названий на углах, а потому слишком уж восторженно оценивает способность чаек к ориентированию.
Каким же образом птица отыскивает и узнает свое гнездо?
Место гнезда
Достаточно очень простого эксперимента, чтобы показать, насколько реакции чаек отличаются от наших. Когда мы в отсутствие чайки вынимаем из гнезда все три яйца и помещаем их на самом виду сантиметрах в тридцати от гнезда, чайка, возвращаясь, обычно направляется к пустому гнезду и после некоторых колебаний часто начинает насиживание. Иногда она поглядывает на яйца перед собой или даже закатывает одно из них в гнездо, но, как правило, на таком расстоянии она их попросту игнорирует. Таким образом, вопреки нашим ожиданиям птица в первую очередь реагирует не на яйца. Гнездо па привычном месте оказывается для нее куда более важным элементом.
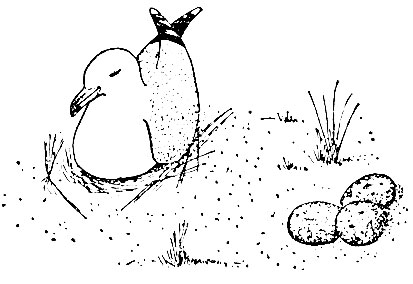
Рис. 18. Чайка, сидящая на пустом гнезде совсем рядом с перемещенной кладкой
Гнездовая ямка
Как воздействуют на птицу гнездо и место, па котором оно расположено? Если удалить пустое гнездо, а ямку засыпать песком, птица, хотя и нерешительно, все-таки садится, как правило, на то место, где раньше было гнездо, показывая тем самым, что место расположения гнезда является существеннейшим элементом в системе раздражителей. Но если под перемещенными яйцами устроить искусственное гнездо, чайка выберет яйца. Следовательно, само гнездо также оказывает на нее воздействие - ведь именно оно склоняет чашу весов в пользу яиц.
Эти опыты не всегда дают одинаковые результаты. Обычно птицы проявляют нерешительность, смотрят на место, где было гнездо, на яйца, а иногда даже присаживаются то на старом месте, то на кладке. Одну птицу может больше привлечь место гнезда, тогда как другая склонна выбирать яйца. Это зависит и от врожденных индивидуал ных особенностей, и от внешних ситуаций, и от ппошлпт опыта птицы, с которой ставится эксперимент Нагшимеп у нас имеются многочисленные данные, свидетельствую щие, что в однообразном окружении - скажем, на травянистой равнине - птицу легче переманить от места гнезда к перемещенной кладке, чем в местности с броскими ориентирами, вроде кустов или неровностей рельефа. Прошлый опыт птицы также немаловажен: птица, насиживающая третью неделю, может быть сильнее привязапа к месту гнезда, чем та, которая только-только приступила к насиживанию. Но как бы то ни было, из этого нельзя сделать вывод, что разнообразие реакций доказывает врожденную способность к выбору. Такой вывод был бы оправдан только в том случае если бы были изучены и различия в индивидуальном опыте, и различия в окружающих условиях.
Эксперименты, которые мы повторяли много раз, ясно указывают, что в систему раздражителей входят как место расположения гнезда, так и само гнездо. Тот факт, что птицы иногда реагировали и на кладку, а также результаты опытов, которые будут описаны ниже, позволяют нам заключить, что яйца также входят в число раздражителей.
Опытов по узнаванию места гнезда мы почти не ставили. Тип местности, в которой обычно гнездятся чайки мало подходит для подобных экспериментов. Смещение предполагаемых ориентиров часто оказывается затруднительным. Тем не менее мы проделали несколько опытов с чайками, гнездившимися на голом берегу, где был какой-нибудь один очень заметный ориентир, вроде ящика или бочонка, выброшенного на песок зимним штормом. Перемещение ориентира в таких случаях мешало чайке найти гнездо. Однако, если гнездо и кладка оставлялись на прежнем месте, замешательство птицы продолжалось недолго - после некоторых колебаний она неизменно находила гнездо. В дюнах, где растительность обеспечивает большое исло ориентиров, чайки, безусловно, используют многие из них. Об этом свидетельствует тот факт, что даже незначительные изменения поблизости от гнезда, несомненно, называются замеченными и вначале мешают чайке ориентироваться, но по-видимому, благодаря тому, что все тальные ориентиры сохранились) чайка всегда находит гнездо. Подобные реакции в очень сложной ситуации, когда в обстановке изменяются лишь незначительные детали, производят большое впечатление, а отличие их от других реакций, когда возможных "сигнальных стимулов" немного, просто поразительно. Как указывает Лоренц [79], способность реагировать на очень сложную ситуацию может быть типична для приобретенных реакций, тогда как для врожденного поведения правилом, по-видимому, является способность реагировать на малое количество сигнальных стимулов. Факты, установленные для серебристой чайки, подтверждают это. Знание примет окружающей местности может быть приобретено только в результате внешних воздействий. Как мы увидим далее, в реакциях на яйца моментов, свидетельствующих о научении, гораздо меньше и на самом деле эти реакции зависят от нескольких простых стимулов. В главе 22 будет приведен еще один разительный пример врожденной реакции на сигнальные стимулы.
"Голубые" яйца
Хотя мы не ставили систематических экспериментов, чтобы выяснить, как чайка находит место гнезда, мы потратили довольно много времени на анализ "узнавания яиц". Если узнавание гнезда было в первую очередь узнаванием места, где оно находилось, то большое разнообразие кладок, особенно в окраске, естественно, заставило нас предположить, что уж собственные яйца птица должна знать. Во всяком случае, это представлялось верным по отношению к птицам, чьи яйца значительно отклонялись от нормы. Как известно, некоторые яйца чаек лишены внешнего - зеленовато-коричневого - слоя пигмента и имеют светлую зеленовато-голубую окраску с очень небольшим числом темных крапин или же вовсе без них. Пометив птиц, в чьих гнездах были обнаружены такие яйца, мы и в следующем году нашли голубые яйца в гнездах тех же птиц. Впрочем, ничего другого мы и не ожидали, так как нам было известно, что голубые кладки из года в год обнаруживаются, как правило, в одних и тех же уголках колонии. Иногда мы находили кладки, в которых одно яйцо было голубым, другое - нормальным, а третье - промежуточным. В 1936 году одна из наших меченых самок, кладки которой в 1934 и 1935 годах были нормальными, отложила сначала голубое яйцо, затем промежуточное и, наконец, нормальное.
Но вернемся к пашей проблеме. Мы решили проверить, узнают ли птицы с голубой кладкой свои яйца, если им предоставить выбор между их кладкой и какой-нибудь другой. Если эти птицы научились распознавать свои яйца, они, несомненно, предпочтут их. С другой стороны, представлялось вполне возможным, что они выберут "нормальную" кладку: если опи не научились распознавать цвет своих яиц и если они обладают, врожденной реакцией на "идеальное" чаячье яйцо, не исключалось, что они предпочтут нормальные яйца своим, отклоняющимся от этого идеала.
Опыт по предпочтению проводился следующим образом. Примерно на расстоянии 30 сантиметров от настоящего гнезда устраивались два искусственных (обычно просто неглубокие ямки). Оставив одно яйцо в настоящем гнезде, остальные два мы помещали в одно из искусственных, а во второе клали два нормальных яйца (фото 29). Затем из укрытия мы наблюдали, какое из яиц, находящихся снаружи, будет водворено в настоящее гнездо. Позднее мы перестали оставлять яйца в настоящем гнезде, а переносили всю кладку в искусственное. Во второе искусственное гнездо мы, разумеется, помещали полную нормальную кладку. Обычно чайка, обнаружив, что гнездо пусто, через некоторое время подходила к одному из искусственных гнезд. Мы содействовали этому, засыпая старое гнездо песком. Разумеется, обе кладки часто менялись местами, а иногда приходилось изменять и местоположение искусственных гнезд. Результаты опытов измерялись числом реакций самца и самки, за которыми мы наблюдали из укрытия. Различались три типа реакций: приближение (и), движение намерения, когда птица взъерошивала брюшные перья и подгибала ноги, словно собираясь сесть (дн), и насиживание (п).
Вот результаты:

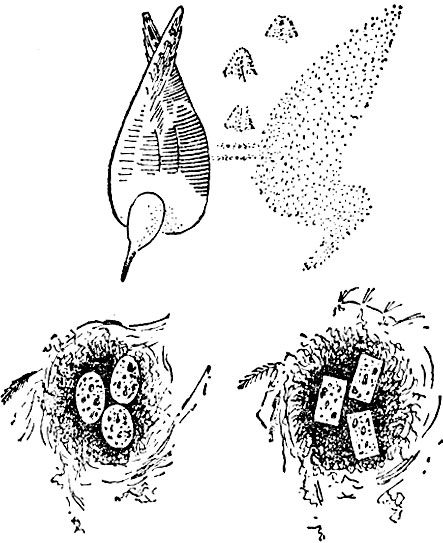
Рис. 19. Опыт с выбором
Эти опыты ставились в течение трех дней (25 и 29 мая и 2 июня 1936 года). Вскоре после 2 июня появились птенцы (точная дата нам неизвестна). Самец проводил в гнезде больше времепи, чем и объясняется полученный с ним более высокий итог.
Как показывают эти цифры, птицы пе делали особых различий между обоими типами яиц, хотя некоторое предпочтение голубым яйцам и имело место. На отсутствие явного предпочтения указывало хотя бы уже нерешительное поведение птиц в каждом опыте. Они не только медлили перед гнездами, поглядывая то на одно, то на другое, но и, подойдя к одному, нередко смотрели на другое, уходил" к нему и даже садились в него, лишь для того, чтобы через какое-то время вновь возвратиться к первому.
Подобный результат оказался несколько неожиданным. Он не просто показал, что чайки даже в таком крайнем случае не "осознавали" (или почти "не осознавали") особого цвета своих яиц, но и как будто свидетельствовал, что нормальный цвет не играет никакой роли в том врожденном "распознавании" яиц, какое, возможно, свойственно птицам.
Деревянные модели
Мы решили установить, какие особенности окраски, размеров и формы яиц могут играть роль в стимулировании поведения насиживания. Метод оставался прежним: мы предоставляли птицам выбор между двумя комплектами яиц, различавшихся только одним каким-либо свойством. Но в методику пришлось внести небольшие изменения. Если в эксперименте, который был описан выше, мы могли пользоваться настоящими кладками, так как требовавшиеся нам различия окраски встречаются в природе, то, приступая к изучению других цветов, а также размеров и форм, мы уже не могли рассчитывать на то, что сами птицы снабдят нас теми далекими от нормы яйцами, которые были нам нужны, например ярко-красными, или двойного размера, или, наконец, кубическими. А поэтому и прибегли к деревянным моделям.
Для начала нам надо было выяснить, примут ли чайки за настоящее яйцо деревянную модель, если ей будут приданы обычные размеры, форма и окраска яйца серебристой чайки. Модели были предложены трем чайкам, и мы зафиксировали их реакцию на поддельную и собственную кладки. Общий результат был таков:
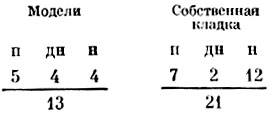
Это указывает на определенное, хотя и не слишком значительное, предпочтение в пользу настоящих яиц. Виновницей такого несоответствия цифр была одна из птиц, которая ни разу пе выбрала модели и 8 раз прореагировала на собственную кладку. Отсюда следует, что остальные две птицы не выказали никакого предпочтения. Но в их поведении наблюдалось качественное различие: реакции па модели чаще ограничивались движением намерения, а настоящие кладки чаще стимулировали насиживание. Общие цифры, однако, невелики, и таких опытов следовало бы поставить гораздо больше. Тем не менее наши деревянные модели, хотя, возможно, и несколько уступавшие настоящим яйцам, служили достаточно хорошей заменой. Во всех последующих опытах мы исключили потенциальные различия между деревянными и настоящими яйцами, предлагая птицам выбирать только между двумя комплектами деревянных яиц.
Воздействие цвета
Опыты проводились с яйцами нормальной окраски, которые сравнивались с ярко-синими, ярко-желтыми и ярко-красными яйцами. На цветные яйца были нанесены как можно более естественные крапины разной величины и разного оттенка, но обязательно того же цвета, что и фон.
Вот результат, полученный в опытах с синими яйцами, которые предлагались пяти разным птицам:

Этот результат, конечно, не был неожиданным. Тем не менее мы невольно удивлялись каждый раз, когда птица безмятежно усаживалась на "пасхальные яйца", как будто всю свою жизнь только это и делала.
В этих опытах, как и во всех других, в которых предлагались два типа кладок, мало различавшихся по стимулирующему воздействию, после нескольких проб (а иногда и после одной) у птицы обычно вырабатывалось четкое предпочтение одному из гнезд независимо от его содержимого - предпочтение месту. В таких случаях мы обязательно перемещали искусственные гнезда вправо и влево от предпочитаемого места, и этого, как правило, оказывалось достаточно, чтобы вынудить птицу выбирать между кладками, а не между их местами. Выбор по месту с такой регулярностью встречался в случаях отсутствия предпочтения той или иной кладке, что постепенно мы начали считать его норным признаком равенства двух типов моделей, которые использовались в опыте. Но когда предпочтения месту не возникало или когда его можно было уничтожить, просто поменяв кладки местами, ото ясно указывало на неравенство воздействия кладок.
С желтыми яйцами мы получили примерно те же результаты, что и с синими. Они предлагались двум птицам.
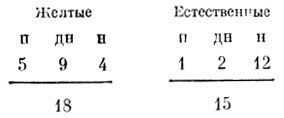
В этих опытах настоящая реакция насиживания также чаще вызывалась естественно окрашенными яйцами, а пе желтыми, из чего, возможно, следует, что птицы как-то их различали.
С красными яйцами картипа получилась совсем другая. Они предлагались семи птицам, и вот с какими результатами:
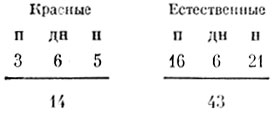
Следовательно, красные яйца оказались гораздо менее привлекательными, чем естественно окрашенные. Предпочтение места проявлялось редко, и его было нетрудно разрушить, переложив па предпочитаемое место красные яйца. Кроме того, мы четыре раза наблюдали, как птица, подошедшая к красным яйцам и, судя по всему, даже собиравшаяся сесть на пих, внезапно принималась их клевать! Это согласуется с наблюдениями Кулемаина [17] и Гете [40]. Оба они сообщали, что чайки, обнаружив в гнезде красные предметы, тут же их выбрасывали. Дирксеп [27] упоминает о сходной реакции у пестроносной крачки. Не исключено, что ото реакция па раненого птенца.
Особенно интересно, что красное яйцо не всегда стимулирует только клевание: порой птица от клевков мгновенно переходит к движениям намерения, связанным с насиживанием. Она, несомненно, разрывается между двумя совершенно разными реакциями. По моему мнению, мы знаем причину этого двойственного поведения. Как мы увидим далее, очень важным раздражителем является форма яйца. Красные же яйца обладали оптимальной формой, но отталкивающим цветом. Красный цвет стимулировал клевки, а округлая форма стимулировала насиживание. Результатом явилась своеобразная смесь двух форм поведения. Это классический тип инстинктивной реакции. Человек в подобной ситуации обдумает противоречивые свойства яйца, решит, считать ли этот предмет яйцом или нет, и поступит соответствующим образом. Не берусь утверждать, что люди ведут себя столь разумно в любой ситуации, но для них это не такая уж редкость. Инстинктивное же поведение - это совсем другое: оно представляет собой непосредственную реакцию на непосредственные раздражители и опирается па гораздо более жесткий механизм, в котором раздражитель и реакция на него связаны очень тесно.
Итак, мы можем сказать, что настоящие яйца оказывают более сильное стимулирующее воздействие, чем самые лучшие деревянные модели, и что различные цвета обладают примерно одинаковым стимулирующим воздействием, хотя некоторые данные указывают, что оно несколько выше у естественного цвета по сравнению с синим и желтым. Стимулирующее же воздействие красного цвета гораздо слабее и даже вызывает совсем иную реакцию.
Интересно сравнить эти результаты с результатами, которые получили другие исследователи. Гете [40] и Штейнбахер 1110] изучали реакции на неестественно окрашенные яйца. Однако их методика отличалась от нашей. Если мы предлагали птице выбор, то и Гете и Штейнбахер изучали реакцию чаек на один комплект яиц, подкладывавшийся в гнездо. Штейнбахер просто сообщает, что птица принимала яйца "коричневато-красные" и черные. Гете подменял настоящую кладку яйцами пеганки (белыми), бледно-голубыми яйцами серебристой чайки и гипсовыми моделями ярко-желтого, зеленого, синего и красного цветов. Он сообщает, что, хотя все яйца, кроме красных, обычно принимались, по поведению птиц было ясно, что они заметили подмену. Кроме того, он упоминает, что птицы клевали красные яйца, а в одном случае - и белое яйцо. Судя по сообщению Гете, его методика меньше подходит для сравнения стимулирующих свойств различных заменителей кладки, чем наша.
Воздействие крапин
Затем мы занялись узором яиц. Если убрать многочисленные темные крапины, которые характерны для всех яиц серебристой чайки, скажется ли это на поведении птиц? Мы изготовили пять комплектов яиц без узоров - белый, черный и различных оттенков серого (светлый, средний и темный). Результаты были следующие:

В результатах не наблюдается особой закономерности. Если белые, серые и черные яйца привлекали явно меньше внимания, чем естественные, то светло-серые и темно-серые по стимулирующему воздействию как будто не уступали естественным. Совершенно ясно, что эти эксперименты следовало бы продолжить. Однако различные результаты можно объяснить и тем, что птица с сильным побуждением к насиживанию менее разборчива, чем та, у которой это побуждение относительно слабое. У нас создалось впечатление, что чайки, как правило, более разборчивы в начале брачного сезона, чем в конце, когда побуждение к насижиь ванию особенно интенсивно. Гете придерживается того же мнения. Кроме того, профессор Бэрендс из Гронингена провел аналогичные опыты уже после нас, пользуясь нашими моделями, и хотя его результаты опираются на сравнительно небольшое число опытов, все его птицы оказались более разборчивыми, чем наши. Од вел наблюдения в колонии, где гнезда постоянно грабились, и, возможно, из-за этого побуждение к насиживанию у его птиц было менее интенсивным, чем у наших. Кроме того, отсутствие строгой закономерности в наших результатах может объясняться еще и тем, что у птиц, с которыми мы проводили опыты, побуждение к насиживанию заметно различалось по интенсивности.
Я думаю, мы вправе сделать вывод, что крапины на яйцах вопреки нашему прежнему мнению [130] играют определенную, хотя и второстепенную, роль и что это особенно заметно, когда стремление к насиживанию малоинтенсивио.
Воздействие формы
Затем мы исследовали воздействие формы. И в этом случае число экспериментов было недостаточным, но первые результаты все-таки кое о чем говорят. Исходя из предположения, что округлость яйца может играть важпую роль, мы изготовили серию моделей с разной степенью закругленности. Один тип представлял собой параллелепипед величиной примерно с яйцо серебристой чайки (фото 28), второй - цилиндр примерно тех же размеров, а третий имел форму призмы. У одних моделей грани были оставлены острыми, у других закруглены. Подробных результатов я приводить не стану, сообщу лишь общие выводы. Параллелепипеды оказались менее привлекательными, чем яйцевидные модели, особенно если грани были острыми (острые грани против яйцевидной формы - 5 : 17, закругленные против яйцевидной формы (9:18). Цилиндры с острыми гранями сильно уступали (8: 18) цилиндрам с закругленными гранями (8 : 11). Призмы не уступали яйцевидным моделям, даже когда грани были острыми (18:12) закругленные против яйцевидной формы - 22:20).
Эксперименты с "острогранными" яйцами дали еще один интересный результат: хотя выбирались они относительно часто, птицы их не насиживали. Если чайка садилась на такие яйца, то долго их переворачивала и затем либо шла к другой кладке, либо все время пыталась найти более удобное положение. Это беспокойство появлялось только после того, как птица прикасалась к яйцам телом, и совершенно очевидно, что оно вызывалось не зрительными, а осязательными раздражителями. Нам стало ясно, насколько неверно истолковывать наши опыты с выбором как свидетельство того, что чайки "узнают" или "не узнают" яйца. Ведь если птицы, сделав выбор, шли к прямоугольным чуркам, значит, они "узнавали" их как яйца. Но когда они оставляли эти чурки, едва к ним прикоснувшись, можно было с тем же основанием сказать, что они "не узнают" их как яйца. Беда в том, что такая терминология подходит для человеческих реакций, но не для реакций птиц. Мы, люди, "узнаем" яйцо, а потом, осмыслив это узнавание рационально, считаем его яйцом. Но птица реагирует по-другому. В этом одна из причин, почему мы иногда достигаем лучших результатов, когда избегаем антропоморфических терминов и описываем поведение птицы через ее реакции на стимулы, получаемые ею от яйца.
Реакции, о которых идет речь, совершенно несомненно складываются в определенную систему; иными словами, насиживание состоит из цепи последовательных действий. Когда серебристая чайка во время поисков корма начинает ощущать побуждение к насиживанию, она возвращается на свою территорию, руководствуясь при этом ориентирами, на которые научилась реагировать. В пределах территории ее путь от сторожевого поста до гнезда тоже определяется знакомыми ориентирами. Приблизившись к гнезду, она начинает реагировать уже на него, и только после того, как партнер встал, на вернувшуюся птицу начинают воздействовать сами яйца, стимулируя стремление сесть на них.
Последняя реакция, как говорят наши опыты, является зрительной, и в ней играют свою роль и местонахождение гнезда, и гнездовая ямка, и кладка. Стимулы, поступающие от яиц, мало связаны с их окраской, но в какой-то мере зависят от крапин и в очень значительной - от округлой формы. Затем в дело вступают осязательные раздражители. Если яйца лежат неудобно, птица реагирует на ото тем, что начинает их переворачивать. Если же не помогает даже многократное переворачивание, птица покидает гнездо.
Наши опыты с формой яиц были довольно неполными но о воздействии формы мы знаем кое-что и помимо них Прежде всего, мелкие неровности на закругленной поверхности яйца всегда вызывают разведывательное поклевывание. И бугорки, и вдавленное™ обязательно обрабатываются клювом. Это очень интересно, так как позже, когда птенец начинает пробивать скорлупу, родители наклевок не трогают. Это объясняется изменением внутреннего состояния птицы: если дать наклюнутое яйцо чайке, насиживавшей не больше недели, она начинает клевать отверстие и, отламывая кусочки скорлупы, может убить птенца
О важности формы свидетельствует еще и следующее обстоятельство. Когда яйцо разбивается (если, например чужая чайка успела его клюнуть), птица перестает его насиживать и съедает (фото 26, 27). Теперь нам понятно почему разбитое яйцо не стимулирует насиживания - оно утратило свой главный раздражитель, свою круглую форму Содержимое яйца, возможно, обеспечивает дополнительный раздражитель, стимулирующий потребность есть И вот поведение мгновенно меняется - птица не насиживает а ест.
Меняя пропорции яиц, но при этом оставляя их округ лыми, мы получали другие результаты. Модели вдвое короче нормального яйца, но нормальной толщины по привле кательности практически не уступали моделям нормальной формы (20 : 24). Однако яйца нормальной длины, но вдвое уже уступали нормальным, и значительно (15 : 34).
Воздействие размеров
И наконец, мы поставили песколько опытов с яйцами нормальной формы, но необычных размеров. Яйца небольших размеров уступают нормальным - те, что были вдвое меньше, вообще не выбирались (0: 13). Очень своеобразные результаты давали яйца, вдвое большие по линейным размерам, то есть в восемь раз превосходившие нормальные по объему.
Когда такую модель положили в одно из искусственных гнезд, а второе гнездо получило нормальную модель, гигантское яйцо было выбрано 6 раз, нормальное же - ни разу. У нас сложилось впечатление, что такое большое яйцо воздействует на птицу гораздо сильнее нормального. При виде большого яйца все птицы приходили в крайнее возбуждение и предпринимали отчаянные попытки прикрыть его своим телом. При этом они неизбежно теряли равновесие, и их маневры, должен признаться, были чрезвычайно забавными.
Эту работу, несомненно, имело бы смысл продолжить. Полученные нами результаты фрагментарны, а нередко и плохо согласуются между собой. Отчасти это объясняется тем, что мы довольно быстро оставляли одну серию епытов и переходили к следующей, не успев получить четких результатов. У нас, правда, есть некоторое оправдание: то, что серии наших опытов слишком коротки, мы поняли лишь тогда, когда работа была уже наполовину проделана. Кроме того, мы избегали слишком часто предлагать одни и те же модели одним и тем же птицам, а поскольку для работы мы, естественно, выбирали птиц с удобно расположенными гнездами или тех, у кого побуждение к бегству было малоинтенсивным, нам приходилось часто менять модели.
Кроме того, наши результаты показывают, что имело бы смысл как-то измерять или по-иному оценивать интенсивность побуждения к насиживанию у каждой отдельной птицы, а также прослеживать процессы научения, возможно, возникающие на протяжении сезона. Следовало бы продолжить опыты с гигантскими яйцами, так как вполне возможно, что они действительно воздействуют сильнее, чем даже настоящие. Это было установлено для кулика-сороки. Судя по всему, кулик-сорока - птица, томящаяся вечной неудовлетворенностью: наши эксперименты доказали не только то, что она обожает насиживать гигантские яйца, а не собственные скромные изделия, по и то, что она с куда большей охотой садится на кладку из пяти яиц, чем на обычную из трех. Ниже я покажу, что подобные "сверхнормальные" раздражители обнаруживались и в других случаях.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'