Стимулы перелетов
При изложении наших знаний о причинах перелетов птиц мы можем начать с того, что уже говорилось в гл. II. Тем самым мы установим связь между предпосылками всей проблемы и частным вопросом о происхождении "перелетного" инстинкта.
Известно, что решающее значение для всех жизненных функций организма имеют железы внутренней секреции и их продукты - гормоны. Выделяемые в кровь в ответ на внутренние или внешние раздражения, они влияют на нервную систему и функции различных органов. Следовательно, от них зависит общее состояние организма. Иногда достаточно очень незначительного стимула, чтобы нарушить гормональное равновесие. Такое же воздействие могут оказать и психические факторы. В результате происходит перестройка всего обмена веществ и изменение инстинктивного поведения животных.
Нам известно, что многие птицы приступают к размножению непосредственно после весеннего перелета, даже при неблагоприятных внешних условиях. Мы знаем также, что смена оперения у перелетных птиц часто связана с началом осенних миграций (рано отлетающие птицы линяют на зимовках, поздно отлетающие - на родине). Отмечено, что птицы задерживаются на гнездовьях, если ко времени отлета молодые птицы еще не научились летать. В аналогичных случаях другие птицы (в частности ласточки) покидали свой выводок. Все эти факты указывают на внутреннюю связь между перелетным инстинктом, инстинктом заботы о потомстве и линькой. Эти процессы, регулярно сменяя друг друга, определяют повадки птиц.
Насколько сильна может быть зависимость одного процесса от другого, особенно наглядно видно на примере жулана: старые птицы отлетают раньше и линяют на зимовке, тогда как молодые меняют мелкие перья до отлета и поэтому позже приступают к миграции. Линька требует больших затрат энергии, и птицы в этот период очень истощены; для перелета они должны вновь накопить известные энергетические запасы. У тех птиц, у которых перелетов нет, например у поганки, согласно Крааку и Хоогерхайде (1942), возможны перелеты, непосредственно связанные с линькой ("Mauserzug"). Подобные миграции наблюдаются у взрослых уток, в большом числе отлетающих в начале августа от берегов Северного моря и Южной Швеции в Гельголандской бухте, где они меняют свои маховые перья; затем эти птицы вновь возвращаются в гнездовую область. Такую линьку, вероятно, следует рассматривать как одну из форм "промежуточного перелета", с той только разницей, что за ней не следует отлета птиц. Даанье и другие авторы доказали, что некоторые виды птиц, например чибисы, свиристели и ряд хищных птиц, могут линять во время осеннего перелета. Линька совершается во время миграции и у буревестников Oceanites oceanicus, которые уже упоминались как летающие птицы в наших широтах. Приведенные данные еще раз показывают, насколько разнообразна действительность, вновь и вновь нарушающая схему общего "правила".
Исходя из связи между возвращением птиц весной с начинающимся вслед за этим размножением, американский исследователь Роуэн (1926, 1929) пришел к выводу, что между перелетным инстинктом и состоянием половых желез, по-видимому, существует тесная зависимость; последние во время перелетов развиты очень слабо и, напротив, в период размножения имеют максимальное развитие. В опытах над серой овсянкой (Junco hiematis) Роуэну удалось доказать влияние солнечной энергии на рост половых желез. Длительное облучение искусственным светом вызвало развитие, а затемнение - инволюцию этих органов. Когда половые железы достигли известного уровня развития, у птицы пробудился перелетный инстинкт, выразившийся в том, что она покинула место эксперимента. Поэтому Роуэн предположил, что причиной проявления перелетного инстинкта является секреция половых желез. Но так как на последнюю влияет количество ежедневно поступающего света, он был склонен считать увеличение или уменьшение светового дня решающим фактором в возникновении перелетного инстинкта. Другой американский исследователь Биссоннетт (1933), работая со скворцами в том же плане, что и Роуэн, также получил аналогичные результаты. Вскоре после этого (1933) Шильдмахер на орнитологической станции Гельголанд продолжил работу Роуэна. Он пытался доказать зависимость перелетного инстинкта от гормонов половых желез, инъицируя самкам горихвосток во время осеннего перелета препарат "прогинон". Применяя определенные дозы этого препарата, ему удалось ослабить или даже погасить проявляемое птицами по ночам перелетное беспокойство. Последнее постоянно регистрировалось при помощи особого прибора. Полученные результаты позволили Шильдмахеру сделать вывод, что осенью проявление перелетного инстинкта является следствием уменьшения количества гормонов, выделяемых половыми железами. Гирсберг и Штади (1934), напротив, пытались возбудить перелетный инстинкт гормональным воздействием. Они воздействовали на пребывающих зимой в спокойном состоянии чижей и щеглов фолликулином. В результате у птиц появилось ясно выраженное "перелетное беспокойство", которое авторы объяснили стимуляцией фолликулином выделения гормонов половой железой, в свою очередь вызвавших проявление перелетного инстинкта.
Здесь мы должны дать краткое объяснение понятия "перелетное беспокойство" и описание аппаратуры, при помощи которой оно регистрируется. Ранее уже упоминалось о том, что содержащиеся в неволе перелетные птицы, когда другие особи их вида приступают к миграции, начинают проявлять беспокойство даже при наилучшем уходе. Они машут крыльями и носятся по клетке, как бы имитируя миграцию, т. е. удовлетворяют свой перелетный инстинкт. Такое явление наблюдается у большинства мелких певчих птиц (насекомоядных) преимущественно ночью, так как на воле их перелет совершается ночью. В опытах это перелетное беспокойство фиксируется при помощи так называемых регистрационных клеток. Каждый прыжок и каждое движение птицы от одной жердочки к другой замыкает электрическую цепь, которая соединена с часовым механизмом, регистрирующим каждое замыкание и размыкание электрической цепи на передвигающейся бумажной ленте (как в аппарате Морзе). По этим записям можно затем судить о длительности и силе прыжков и беспокойства (рис. 42). Однако теории Роуэна и Шильдмахера вскоре подверглись критике. Прежде всего, они относятся только к тем видам птиц, которые не перелетают из северного полушария в южное, где укорочение светового дня сменяется его удлинением, но тем не менее половые железы зимующих там птиц не развиваются. Кроме того, этим взглядам противоречит поведение уток, половые железы которых сильно развиты зимой, следовательно, задолго до начала весеннего перелета. Указанные теории не могут объяснить перелеты молодых птиц с еще совершенно неразвитыми половыми железами. Наконец, подобные широкие обобщения нельзя делать из-за небольшого числа положительных результатов по сравнению с количеством отрицательных данных. Итог этим соображениям подвел сам Роуэн (1932) опытами над воронами. У одной группы подопытных птиц он удалил половые железы, другую облучал светом, а третьей инъицировал гормон. В результате оказалось, что поведение всех этих птиц по существу мало чем отличалось от поведения контрольных птиц, которые не подвергались никакому воздействию. Ряд дальнейших опытов, проведенных Шильдмахером, а затем и Путцигом (1937-1939) на орнитологической станции Росситтен, показал, что освещение многочисленных мелких птиц зимой хотя и вызвало развитие половых желез, но не привело к преждевременному проявлению перелетного беспокойства. И наоборот, летом птицы в регистрационных клетках часто находились в состоянии перелетного беспокойства до тех пор, пока оно не прерывалось линькой. В соответствии с результатами Роуэна Путциг установил, что как весной, так и осенью выпущенные на волю кастрированные перелетные птицы (клуши и обыкновенные чайки) вели себя точно так же, как и физиологически вполне нормальные представители того же вида. Сходное поведение наблюдалось также у ворон и галок, которым был введен половой гормон самок. Ган (1939) проводил аналогичные опыты с североамериканскими певчими птицами и получил такой же результат.

Рис. 42. Прибор для регистрации перелетного беспокойства птиц на орнитологической станции Гельголанд (в дальнейшем он был усовершенствован)
Таким образом, было установлено, что в противоположность первоначально распространенным взглядам инстинкты спаривания и насиживания, обусловленные воздействием гормонов половых желез, никоим образом не следует приравнивать к инстинкту весеннего перелета. Первые могут рассматриваться только как дополнительный фактор, способный ускорить или прекратить перелет. Что касается инстинкта осеннего перелета, то он не находится в прямой зависимости от гормона половых желез.
После первых поразительных результатов в этой области исследователи стали предостерегать против возможной переоценки единичных данных. Учитывая видовые различия в поведении подопытных птиц, необходимо избегать всякого рода обобщений. Тем не менее весьма вероятно, что приведенные выше данные о зависимости между развитием половых желез и миграцией относятся ко всем перелетным птицам.
При обсуждении связи между перелетами птиц и погодой мы указывали на отличия птиц, перелет которых определяется внешними метеорологическими условиями, от птиц, миграции которых зависят от инстинкта. При этом мы отмечали, что в противоположность первоначальному пониманию этих терминов даже типичные "инстинктивные" птицы в известных условиях ведут себя как "погодные" птицы, так как у представителей обеих групп миграции могут вызываться воздействиями внешней среды.
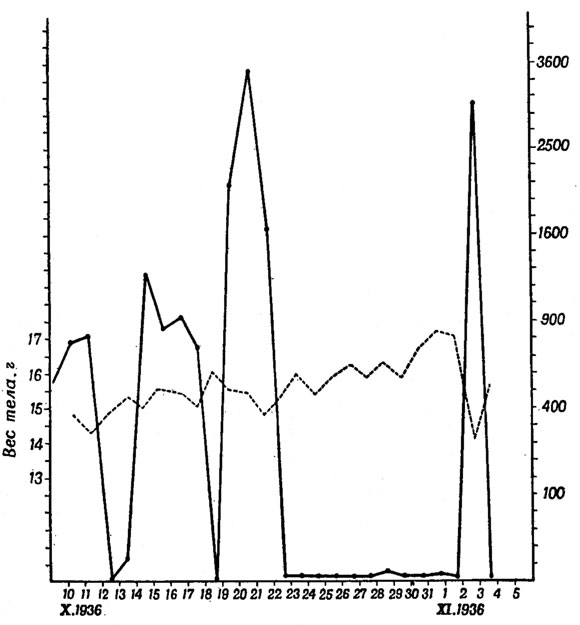
Рис. 43. Зависимость между перелетным беспокойством и весом тела у серой славки (Sylvia с. communis Lath.) (Из Меркеля, 1940.) — перелетное беспокойство; --- вес тела
Влияние таких факторов на птиц первой группы сказывается и в том, что у птиц, залетевших в Германию из чужих стран, часто в связи с приспособлением к местным климатическим условиям сдвигаются периоды размножения. Германские птицы, перевезенные в тропические области или южное полушарие, меняют время перелета или же вообще перестают мигрировать. Кроме того, отмечено, что содержавшиеся в течение многих лет в клетках перелетные птицы постепенно переставали проявлять беспокойство в период перелета. Характерно, что это чаще всего наблюдалось у таких видов, которые на воле склонны к оседлости. Выше мы уже неоднократно упоминали эти виды в связи с другими вопросами.
В этом отношении представляет интерес установленный Томасом (1934) и Хиксом (1938) факт, что европейские скворцы вели себя в США в течение нескольких десятилетий, как оседлые птицы, но в последнее время стали совершать перелеты, направления которых соответствовали направлениям пролетов европейских скворцов. Переведенные в Шотландию канадские казарки - типичные перелетные птицы - превратились в оседлых; птицы, выращенные Бенгт Бергом в окрестностях Кальмара (Швеция), улетели вместе с серыми гусями, следуя вдоль берегов Центральной и Западной Европы. Переселенные из Англии в Финляндию кряквы приспособились к поведению местных птиц этого вида.
Но если "погодные" птицы всегда отвечают на внешнее раздражение началом миграции, то, как уже упоминалось, "инстинктивные" птицы приступают к перелету только при наличии особых предпосылок. В некоторых случаях внешнее влияние оказывается неэффективным и непосредственной причиной отлета являются сдвиги гормонального равновесия, - влияющие на обмен веществ. Таким образом, мы переходим к физиологически обоснованным различиям между этими группами птиц и к оценке значения физиологии обмена веществ для изучения перелетов вообще.
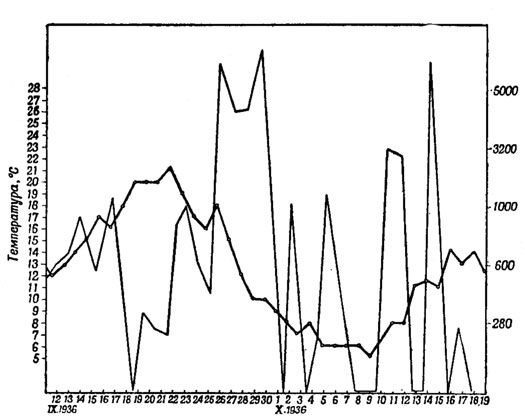
Рис. 44. Зависимость между температурой и перелетным беспокойством у серой славки (Sylvia с. communis Lath.). (Из Меркеля 1940.) О-О температура; ------ перелетное беспокойство
Первым ученым, положившим начало такого рода исследованиям, был Греббельс. В своем большом труде "Птица" (1932) он изучал зависимость между питанием птицы и ее перелетом и исследовал значение усиленного отложения жира в период перелета, которое он рассматривал как физиологический эффект кастрации. В связи с этим он собирал данные о весе птиц, определял состояние питания мигрирующих птиц, их пищеварение и температуру тела, чтобы таким путем получить представление о связях между обменом веществ и перелетом. Приблизительно в то же самое время Вагнер (1930) изучал зависимость суточного ритма перелетных птиц от внешних факторов. В дальнейшем (1937) он установил, что у типичных перелетных птиц количество пищи оказывало небольшое влияние на возникновение перелетного беспокойства, а у менее выраженных перелетных птиц это влияние было очень значительным. Высокая температура оказывала задерживающее действие, низкая - способствовала проявлению ночного беспокойства. Меркель (1937) принял терминологию Греббельса о "предрасположении" и "готовности" к перелету и на основании ряда собственных опытов пришел к выводу, что предрасположение к перелету, по существу, идентично способности накапливать жир, а перелетное настроение равнозначно началу перелетного беспокойства и вызывается небольшим повышением секреции щитовидной железы, вследствие чего мобилизуется резервный обмен веществ (см. гл. III). Следовательно, при наличии готовности к полету небольшое понижение температуры явилось бы (через повышение секреции щитовидной железы) непосредственной причиной отлета.
Американские исследователи в опытах над голубями, домовыми воробьями и крапивниками получили иные результаты. Они обнаружили видовые отклонения, явившиеся следствием различий в характере обмена веществ у разных видов. В то время как гуси и утки оставались совершенно спокойными в течение всего периода голодания и тем самым понижали свой обмен веществ до минимума, волнистые попугайчики и канарейки пытались поддержать температуру тела при помощи движений, за счет запасов жира и гликогена. Согласно Путцигу (1938а), этот факт указывает на значительные различия в обмене веществ и в связи с этим также и в механизме терморегуляции "погодных" и "инстинктивных" птиц. Последние - к ним относятся в первую очередь насекомоядные птицы - "должны при более быстром переваривании и значительном расходе кислорода получать гораздо больше пищи и много двигаться, чтобы поддержать свою в норме высокую температуру тела". В условиях укорачивающегося дня в конце лета эти птицы не могут добывать необходимое для поддержания процессов окисления количество пищи, особенно когда первые холодные ночи ускоряют сгорание жира и гликогена. В результате появляется "психическое" беспокойство, птицы начинают усиленно двигаться, что соответствует началу истинного перелета.
Однако последние работы Путцига (1939) показывают, насколько мало разработаны эти вопросы и сколь ошибочно было бы переносить выводы о причинах поведения одного вида птиц на другой. В этих работах Путциг в противоположность своим прежним предположениям и данным других авторов пришел к выводу, что, несмотря на то, что накопление жира способствует перелету, а его отсутствие задерживает миграции, птицы иногда мигрируют и с минимальными энергетическими запасами. Во всяком случае, у скворцов, зарянок и горихвосток проявление перелетного инстинкта не зависит от их физиологической конституции. Возможно, это относится и к другим видам, однако точных данных об указанной закономерности пока нет. Вне зависимости от того, будут ли получены положительные или отрицательные результаты, мы приближаемся к разрешению очень важного вопроса о взаимосвязи перелетов и процессов обмена веществ. Путциг предложил заменить старые понятия "Wettervogel" и "Instinktvogel", которые он считает слишком односторонними и в некоторых отношениях вводящими в заблуждение, более нейтральными определениями: "птицы, перелеты которых обусловлены внешней средой, и птицы, перелеты которых обусловлены внутренней средой" ("Aussenwelt- und innenweltbedingte Zug-vögel"). Различие между обеими группами он определил следующим образом: для того чтобы представители первой группы приступили к миграции, необходим внешний толчок, заключающийся в неприятном самочувствии вследствие низких ночных температур в сочетании с длительной темнотой или же вследствие недостатка корма. "В новых условиях у таких птиц обнаруживается непосредственное воздействие пережитого на выполняемые ими движения. У птиц, перелеты которых обусловлены внутренней средой, также возможен подобный толчок к началу перелета и его усиление в результате воздействия внешних раздражений. Последние передаются рефлекторно или же посредством эндокринных желез и обмена веществ. Однако гораздо чаще "пусковой механизм" перелетов этих птиц обусловлен автоматизмом нервной системы. У обеих групп целесообразное поведение наблюдается и при отсутствии опыта, что составляет основную проблему перелетов птиц".
Говоря об обмене веществ и температурном режиме у перелетных птиц, мы неоднократно указывали на роль щитовидной железы, а также коснулись ее значения для начала перелетов. При низкой температуре происходит усиление выделения гормона щитовидной железы (тироксина), который путем повышения интенсивности всех процессов жизнедеятельности организма вызывает расходование запасных веществ. Установлено, что щитовидная железа перелетных птиц выделяет большое количество гормона. Зимой, однако, ее секреция значительно понижена. Исходя их этих ритмичных изменений функции железы, был сделан вывод, что тироксин играет решающую роль для начала перелета. Вагнер (1930) отметил у подопытных птиц признаки перелетного беспокойства после кормления их щитовидной железой. Путем инъекции тироксина Меркель (1937) также вызвал усиление активности у птиц, перелет которых уже был закончен. Проведенные им гистологические исследования показали, однако (в отличие от данных других исследователей, например Кюхлера, 1934), что в период перелетов тироксин выделяется в небольшом количестве. Его содержание особенно велико в период линьки. После опытов Вагнера Греббельс отметил, что повышение функции щитовидной железы во время перелетов невозможно согласовать с понижением основного обмена веществ. Гистологическими исследованиями (Путциг, 1937) установлено, что состояние щитовидной железы у разных видов залетных птиц (клёст, пестрый дятел, свиристель, снегирь, а также чайки, аисты и некоторые хищные птицы) очень изменчиво, и поэтому на основании ее функции нельзя вывести никаких общих заключений относительно перелетов птиц. Ритмичность функции щитовидной железы (как и половой железы и других желез внутренней секреции) иногда находится в противоречии с ее реакцией на раздражения окружающей среды, результат которой в данном случае зависит от общего состояния организма.
Путциг (1938) обращает внимание на то, что усиление функции щитовидной железы отражается не только на обмене веществ, но и на нервной системе, что приводит к повышению чувствительности на раздражения извне. И, наоборот, воздействие центральной нервной системы может вызвать изменения в органах. Этих взглядов придерживается и Штади (1938); он особенно подчеркивает возможность ошибок из-за непосредственного сравнения поведения подопытных птиц, содержащихся в клетках, с поведением птиц, живущих на воле. Содержание птиц в клетках может через центральную нервную систему вызвать усиление функции щитовидной железы. Нормальному протеканию процессов обмена мешает также ограничение подвижности подопытных птиц, что не может не отразиться на функции щитовидной железы. Подобные влияния, разумеется, трудно учесть в эксперименте. Однако возможно, что именно они и являются ключом к пониманию огромного разнообразия явлений, на основе которых возникли обобщения, которые всегда оказывались лишь индивидуальной или же, в лучшем случае, свойственной определенному виду реакцией.
В нескольких работах Путциг указывал на особую роль гипофиза во всей системе желез внутренней секреции, а, следовательно, и в процессе перелета птиц. Он отмечает необходимость изучения гонадотропного гормона передней доли гипофиза, влияющего на секрецию половых желез. В дальнейшем разработкой этою вопроса занялись американские и английские исследователи, которые за последние годы достигли в этой области значительных результатов. Некоторые из них мы здесь вкратце приведем.
Рилей, Гарднер и Уитши (1938) нашли, что гипофиз самок очень слабо реагирует на световые раздражения и что для стимуляции функции гипофиза, а, следовательно, и для созревания яичника очень большое значение имеют "психические" факторы. Бергер, Биссоннетт и Дулиттл (1942) в опытах над скворцами пришли к выводу, что половые железы активирует не свет, а длительное бодрствование. Вольфсон (1941) исследовал роль гипоталамической области промежуточного мозга, которую следует рассматривать как центр регуляции сна. Если гипоталамус не возбуждается зрительными раздражениями, то уменьшается его активность, а следовательно, и активность мозга, так же как и всего организма. Но, поскольку гипоталамус воздействует на гипофиз, длительное световое раздражение (при длинном дне или искусственном освещении в эксперименте) усиливает функцию гипофиза и тем самым - развитие половых желез. Хотя последовательность биологических циклов у птиц - размножения, линьки и перелета - как будто наследственно закреплена, однако проявление каждого из них зависит от внешних раздражений, различных у разных видов и нередко даже у разных особей. Вольфсон (1942) исследовал вопрос о происхождении готовности к перелету. Изучив перелетных и оседлых представителей одного из видов овсянок (Junco oregonus), он установил, что на готовность к перелету влияет непосредственно гипофиз. Его действие заключается в создании определенного физиологического состояния, которое обеспечивает птице необходимую для миграции энергию. В отличие от перелетных птиц у оседлых Вольфсон не обнаружил накопления жира весной. У последних рано активируется функция семенников. Вольфсон особенно подчеркивал важность для регулирования всех видов внутренней секреции "психических" факторов (наряду с такими, как длина дня, температура и пища). На это, как известно, уже указывали Путциг и другие авторы. В итоговой работе Вольфсон (1945) пытался точнее определить, какой гормон влияет на готовность к перелету, однако он не смог получить исчерпывающих результатов. Изучение составных частей комплекса внутренних раздражителей выявило огромное влияние на все функции организма гипофиза, который, собственно, управляет общим физиологическим состоянием, а, следовательно, и поведением животного.
Приведенные данные указывают, что вопрос о значении гипофиза в общем гормональном балансе организма не решен. Подробное изложение влияния гипофиза на перелет птиц выходит за рамки этой книги, и, поскольку указанные вопросы еще только разрабатываются, это может отрицательно сказаться на общем обзоре, который мы предлагаем читателю.
Если мы после анализа физиологических исследований и сравнения полученных результатов с фаунистическими данными хотим еще раз охарактеризовать современный взгляд на сущность перелетов птиц и вызывающего их перелетного инстинкта, то мы должны признать, что привычное мнение о перелете, как о строго определенном, доминирующем и неизменном явлении, может существенно измениться. В настоящее время (во всяком случае, в отношении целого ряда видов, о которых имеется достаточно данных) установлено, что перелеты птиц не определяются факторами наследственности, что находится в известном противоречии с данными, приводившимися при характеристике методов изучения перелетов птиц. Но тогда речь шла только об основном правиле, которое и в настоящее время в известной степени имеет силу. То обстоятельство, что это правило в ряде отношений кажется нарушенным и что в результате развития наших знаний такие нарушения, возможно, будут обнаруживаться и в дальнейшем, служит лишь доказательством изменчивости всех процессов в природе, среди которых и перелеты птиц не являются исключением. "Перелет птиц - это не статика, а динамика",- говорил Путциг в одной из своих последних работ. Он дал следующее определение понятия "перелетного инстинкта": "Перелетный инстинкт - это обусловленный историей вида и физиологическими реакциями организма приобретенный автоматизм, основанный на преобразовании сил внешней среды во внутренние, осуществляющийся непосредственно или же через систему желез внутренней секреции, результат действия которого гарантирует сохранение вида".
На этом мы заканчиваем главу об изучении перелетов птиц, слишком краткую и сжатую по сравнению с ее значением. По решению частных вопросов и по выводам эта глава наименее зрелая, поскольку работы в этой области еще не закончены и окончательных результатов не получено. Это лишь первые попытки проникновения в неизведанные края, куда наука получила доступ лишь недавно. Но за это короткое время сделано так много, что мы вправе надеяться со временем получить ответ и на самый трудный, основной вопрос науки о перелетах птиц: "почему?" Путь к этой цели известен - это методы физиологического исследования, а достижение ее - вопрос времени.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'