Что ежегодно вызывает перелеты?
Итак, мы знаем, для чего нужны птицам перелеты. Оставшиеся птицы попадают зимой в неблагоприятные условия и, как правило, погибают. Большинство перелетных птиц не приспособлено к перенесению суровых зим. Вальгрен изучал в 1955 г. два вида овсянок - перелетную садовую и оседлую обыкновенную. Та и другая летом живут в Финляндии вполне благополучно, но садовая овсянка не смогла бы перезимовать в Финляндии. У нее зимой высокий уровень обмена веществ, невместительный пищеварительный тракт. Чтобы поддерживать свое существование, она нуждается в значительно большем количестве пищи, чем оседлая овсянка. Кроме того, при понижении температуры воздуха у нее сильно возрастают затраты на поддержание температуры тела. В результате всего этого, даже при обилии корма, садовая овсянка должна при температуре воздуха 0° питаться в течение 10 часов в сутки. Обыкновенная овсянка при температуре -10° довольствуется 6-часовой кормежкой. Ясно, что садовая овсянка не может зимовать в Финляндии.
Можно ли предположить, что она улетает каждый год потому, что ее гонит холод? Иными словами, могут ли ежегодный отлет птиц вызывать первичные факторы среды - те факторы, с помощью которых осуществляется естественный отбор? Подобные предположения и ныне приходят на ум каждому, кто начинает раздумывать о возможных стимулах перелетов.
Почему бы не предположить, что общее ухудшение условий существования осенью гонит птицу к югу, а весной, с улучшением условий они возвращаются вновь? Каждый, кто наблюдал птиц в течение нескольких лет, знает, что если весна холодная, то первые перелетные птицы появляются позднее, а если теплая, то раньше. В средней полосе в холодные годы зимует много снегирей, свиристелей, чечеток, а в теплые зимы их может совсем не быть. Многие виды птиц не улетают на зимовку, если осенью большой урожай рябины. Например, урожайной зимой 1964/1965 г. остались зимовать дрозды-рябинники. Большие стаи этих птиц можно было видеть всю зиму под Ленинградом. Немецкие орнитологи назвали таких птиц "погодными мигрантами", т. е. птицами, отлетающими под непосредственным давлением первичных факторов. Погодным мигрантам противостоят "инстинктивные мигранты" - такие виды, которые улетают и прилетают ежегодно в одни и те же сроки, независимо от погодных условий в данном году.
Различия между этими группами действительно существуют, но название это, хотя и удобно, но по существу неудачно. Оно часто приводит к неверному пониманию сути явления. Дело в том, что и у "погодных мигрантов" изменения погоды не во все сезоны года вызывают перелет, а лишь в строго определенные периоды осени и весны. В другие сезоны ухудшение условий может погубить птицу, но на зимовку она не улетит. Значит, и у погодных мигрантов тоже есть миграционное состояние, и только в этом состоянии погодные и пищевые стимулы вынуждают птиц к отлету.
Итак, согласно гипотезе о стимуляции перелетного состояния первичными факторами, оно возникает весной в результате улучшения условий жизни - потепления, стаивания снега, появления пищи, увеличения длины кормового дня. После прилета на гнездовье перелетное состояние прекращается в связи с увеличением затрат организма на гнездование и линьку. Оседлые виды не совершают перелетов, потому что токование и гнездование начинаются у них раньше и поглощают весенний избыток энергии. Осенью, после линьки, затраты организма снижаются и избыток энергии обеспечивает осеннее миграционное состояние. У оседлых видов линька растягивается до похолодания, что мешает отлету. По мнению других специалистов, осенняя миграция, наоборот, результат ухудшения условий.
Чтобы разобраться в этом, нужно строго разграничить внешние факторы на группы: первая группа факторов - исторические первопричины миграции, о них мы говорили выше; вторая - факторы, вызывающие наступление перелетного состояния; и третья - влияющие на реализацию этого состояния. Смешение этих факторов приводит к страшной неразберихе. Приведенная выше гипотеза касается только факторов, вызывающих появление перелетного состояния. Описано много опытов, в которых перелетное состояние пытались вызвать в середине зимы или лета обильным кормлением или, наоборот, недостатком пищи, холодом или теплом, длинным или коротким кормовым днем, - все тщетно. В каких бы условиях мы ни содержали птицу, у нее в положенное время появится перелетное состояние, в положенное время оно и прекратится.
С этими опытами, доказывающими, что перелетное состояние у птиц вызывают не погодные и пищевые факторы, очень часто путают опыты по выяснению третьей группы факторов - так называемых непосредственных стимулов к миграции. Те, кто смешивают эти две группы факторов, допускают серьезную логическую ошибку: они не отличают причину от повода. Непосредственные стимулы - это лишь повод к проявлению перелетного состояния, которое возникает под влиянием второй группы факторов. Есть много сообщений, начиная с работы немецкого орнитолога Шильдмахера, опубликованной в 1938 г., о том, что в конце зимы у некоторых птиц, в частности зарянок, потепление окружающего воздуха вызывает ночное перелетное беспокойство. Осенью тот же результат можно получить при содержании птиц на холоде. Еще раньше, в 1930 г., другой немецкий ученый, Вагнер, установил, что осенью обильное вечернее кормление прекращает ночное беспокойство у дроздов, а недостаток пищи усиливает его. Славки, типичные инстинктивные мигранты, не меняли своего поведения в подобных опытах.
Эти опыты неоднократно были повторены как в Европе, так и в Америке. Оказалось, что весной и осенью все погодные мигранты очень чувствительны к изменениям погоды и пищи. У многих из них начало перелетного беспокойства очень сильно оттягивается, если их содержат осенью в тепле и достатке пищи, а весной на холоде.
Но перелетное беспокойство - финал предмиграционных изменений в организме. Сами же изменения и у погодных мигрантов не зависят от погоды и пищи. Погодные мигранты так же регулярно, как и инстинктивные, приходят в перелетное состояние. Разница между ними лишь в том, что первые, подготовившись к отлету, "ждут" соответствующих погодных условий, а вторые сразу начинают миграцию.
Так, осенью зарянки, уже накопившие жир и готовые к перелету, обычно держатся на одном месте, пока не выдастся ночь с хорошим заморозком. А славки, например, отлетают, не дожидаясь ухудшения погоды. Следовательно, зарянки нуждаются в непосредственном стимуле к миграции со стороны погоды, а славки не нуждаются в нем: для них таким стимулом служит окончание подготовки к миграции. Но и зарянки, если погодный стимул слишком запаздывает, улетают, не дожидаясь его.
Погодные мигранты - совсем не "архаическая ступень эволюции перелетов", как думали раньше. Птицы приспособились как можно дольше задерживаться на гнездовье осенью и как можно раньше возвращаться весной. Это виды, улетающие сравнительно недалеко, поэтому им выгодно улетать лишь тогда, когда плохие условия уже наступили.
Дальние мигранты так поступать не могут. Они отлетают заранее, когда условия жизни вполне удовлетворительные. Далекое, продолжительное и напряженное путешествие нельзя ставить в зависимость от капризов погоды. К тому же эти птицы просто неспособны переносить ухудшение условий - кормиться и поддерживать температуру тела при резком похолодании не могут, так как они приспособлены к нашему лету и теплым зимовкам. Эти птицы так никогда и не встречаются лицом к лицу с теми неблагоприятными условиями, для защиты от которых у них выработаны перелеты. Такая встреча равнозначна их гибели.
Итак, дальние мигранты готовятся к перелетам и улетают до наступления холодов и бескормицы. Ближним мигрантам не так страшна встреча с первыми признаками наступающей зимы. Но и они, подобно дальним мигрантам, готовятся к перелету еще до появления первых холодов. Таким образом, предположение о том, что погодные и пищевые условия ежегодно вызывают перелетное состояние, не подтвердилось в опытах и ныне отвергнуто. Эти стимулы могут лишь ускорить или задержать начало отлета, да и то лишь у ближних мигрантов.
Для того чтобы заранее подготовиться к перелету, необходимо как-то заблаговременно определить приближение времени отлета. Для этого нужен какой-то сигнал из окружающего мира, извещающий каждый год о приближении весны или осени.
Тут следует ввести одно понятие - сигнальный фактор, т. е. событие в окружающем мире, безразличное (не вредное и не полезное) для птицы, но предупреждающее о приближении важного первичного фактора. На поиски такого сигнала учеными было потрачено много сил. Выше уже говорилось, что опыты с содержанием птиц в тепле и на холоде изменяют времени наступления перелетного беспокойства, но никогда не меняют времени наступления перелетного состояния. На первый взгляд это обстоятельство кажется очень странным, но если подумать - оно вполне естественно.
Если бы птицы пользовались изменением температуры окружающего воздуха как сигналом о приближении поры отлета, они постоянно ошибались бы. Подготовка к отлету (предмиграционный период) длится несколько недель. Перелет тоже занимает длительное время - месяц, а то и три. На протяжении всего этого времени погода должна быть благоприятной для жизни птицы: перелет - большая нагрузка. За это время птицы преодолевают огромное расстояние, и если перед отлетом в данном месте тепло, это совсем не значит, что теплая погода будет на всем пути перелета. Если птицы из-за теплой погоды улетят позже обычного, неожиданное похолодание на пути пролета может привести к катастрофе. Горький опыт сотен поколений предков подсказывает птицам, что готовиться к отлету необходимо всегда в одно время.
Непригодность температуры как сигнала для начала подготовки к перелету особенно наглядна на примере весеннего отлета птиц, зимующих в тропиках. В этом случае по изменению погоды на зимовках нельзя судить о состоянии погоды на севере, а тем более о погоде через два месяца, когда птицы достигнут гнездовий. Для птиц, зимующих вдали от гнездовий, особенно в теплом поясе, нужен какой-то иной сигнал. Этот сигнал должен из года в год повторяться в одно и то же время. Есть только один сигнал, удовлетворяющий этим условиям, - длина светлой части суток. По длине дня можно точно установить ту или иную дату везде, кроме экватора.
Предположение о регуляции перелетов изменением продолжительности светлой части суток, или, как говорят ученые, о фотопериодической регуляции, впервые проверил канадский зоолог Роуэн. 1926 г. - дату появления его первого отчета об опытах на американской овсянке - сизом юнко - принято считать началом современных экспериментальных работ по изучению перелета птиц. В последующие годы вышло в свет еще несколько работ этого выдающегося исследователя. В 1937 и 1938 гг. подобные опыты проделали Шильдмахер в Германии, в 1945 г. Волфсон в США, после чего количество сообщений об опытах по регуляции перелетов светом начало расти, как лавина. К нашему времени изучено уже около ста видов птиц.
Роуэн поймал зимой в Канаде сизых юнко, которые там зимуют (гнездятся они на Аляске), и поместил в клетках прямо на открытом воздухе. Одни птицы (контрольные) содержались в клетках при естественной длине дня, а другие - при искусственно продленном с помощью электрического освещения. Невзирая на зимний холод юнко, жившие в условиях продленного дня, начали подготовку к весенней миграции. У птиц, живших в естественной обстановке короткого зимнего дня, состояние не менялось до весны. Затем они в положенные сроки тоже начали подготовку к миграции (рис. 10).
Роуэну, следовательно, удалось спровоцировать юнко к весенней миграции удлинением светового дня. Позднее другие авторы подробно выяснили, как действует свет на птицу. Сначала предполагались три пути воздействия: прямой через глаза, и два косвенных - увеличение продолжительности кормового дня и подвижности. Несколько остроумных опытов показали, что косвенные пути не имеют существенного значения.
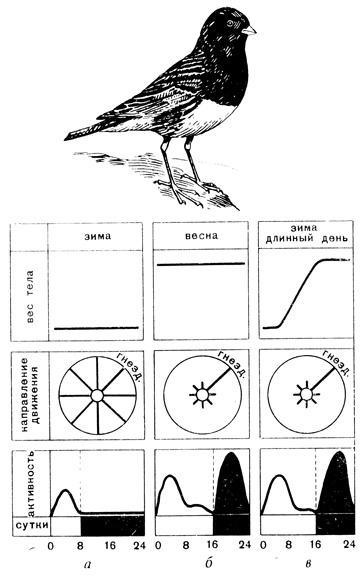
Рис. 10. Опыты с сизыми юнко. а - результаты опытов с зимними птицами при коротком дне, б - во время весенней миграции, в - перелетное состояние вызвано зимой
Наиболее убедителен следующий опыт, проводившийся в Америке и СССР. Одну группу птиц содержали зимой в условиях короткого дня, вторую в условиях длинного, а третью при коротком дне, но на протяжении ночи, через каждый час повторялись короткие вспышки света. Птицы этой группы кормились и двигались столько же часов, сколько и птицы при коротком дне, а перелетное состояние у них появилось одновременно с птицами, жившими при длинном дне. Это объясняется тем, что прерывистое освещение действует так же, как непрерывное: к этому времени уже было известно, что короткая вспышка света "снимает" значительный период темноты. Опыты по изучению фотопериодических реакций стали важным орудием в руках экспериментаторов. С их помощью установлено, что световые сигналы воспринимаются глазом и что поступают они в особый отдел промежуточного мозга - гипоталамус.
В настоящее время многие ученые почти не сомневаются в том, что осенняя миграция также вызывается определенной длиной светлой части суток, но подтвердить это опытами не удается. Возможно, в этом случае регуляция гораздо сложнее. Выяснено, что те виды птиц, которые начинают весеннюю миграцию раньше, реагируют уже на небольшое увеличение длины дня, а виды птиц, поздно начинающие миграцию, - только на длинный день (14-15 часов света в сутки). Даже в пределах одного вида северным популяциям для начала весенней подготовки к миграции необходим более длинный день, чем южным. В пределах одной популяции самки часто менее чувствительны к сигналу, чем самцы, поэтому самки начинают весеннюю миграцию позднее.
Длина светового дня - типичный пример так называемого сигнального фактора. Само по себе увеличение длины дня на несколько минут безразлично для птицы, так как почти ничего не изменяет в окружающем мире. Но оно служит сигналом приближающихся изменений в природе, и этого вполне достаточно, чтобы в процессе эволюции у птиц выработалась реакция на этот сигнал, которая выражается в подготовке к миграции.
Для птиц, зимующих в северном полушарии, сигналом для начала отлета на родину служит увеличение длины дня. Птицам, зимующим в южном полушарии, сигналом для отлета тоже служит изменение длины дня: день там весной сокращается. Как определяют время отлета птицы, зимующие на экваторе, где продолжительность дня и ночи всегда одинакова? Первоначально это было основанием для возражений против взглядов Роуэна. Но теперь известно, что после окончания весеннего перелета в организме птицы наступает особое состояние - рефрактерность. В этом состоянии механизмы, включающие весеннюю миграцию, не действуют. Для того чтобы способность реагировать на свет восстановилась, нужен короткий день. Это состояние регулируется длиной светового дня: она тем продолжительнее, чем длиннее день, и тем короче, чем короче день. Птицы, зимующие севернее экватора, теряют рефрактерность обычно еще в ноябре - начале декабря. А у птиц, зимующих на экваторе, рефрактерность из-за более длинного дня заканчивается только к весне. Таким образом, продолжительность рефрактерной фазы строго зависит от длины дня: так как начало ее на гнездовье точно определено длиной дня, то и окончание фазы на экваторе произойдет в определенное время. Часто такое явление называют внутренним ритмом организма. На самом деле этот внутренний ритм, конечно, контролируется внешним сигналом: если бы день на экваторе стал вдруг не 121/2 часов, а меньше, рефрактерность закончилась бы раньше и миграция началась бы раньше. Опыты с птицами, улетающими на экватор, подтвердили это предположение.
Итак, в большинстве случаев перелет начинается не под воздействием изменения важных для птицы условий жизни, а под влиянием предупреждающего сигнала. Даже в тех случаях, когда момент отлета зависит от состояния погоды и обилия пищи, подготовка к отлету происходит задолго до этого и всегда под контролем сигнального фактора. Для подавляющего большинства наших птиц таким сигнальным фактором служит продолжительность светлой части суток - единственный сигнал, который может повсеместно дать представление о времени года.
В дальнейшем будет идти речь о некоторых тропических птицах, для которых таким сигналом служит не длина светлой части суток, а изменение влажности.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'