Глава 14. Происхождение ритуального демонстративного поведения при ухаживании
Вздергивание головы
Необходимо сказать несколько слов о вздергивании головы как о предкопуляционном демонстративном поведении. Самка точно так же вздергивает голову, когда выпрашивает корм. Похожим, но несколько иным движением выпрашивают корм взрослые птенцы. Различия в основном проявляются в голосе: молодая серебристая чайка испускает не очень громкий хриплый писк, а выпрашивающий крик взрослой птицы отличается тем ясным чистым тембром, который вообще характерен для большинства криков серебристых чаек. В нем слышится определенная нежность, которой нет в мяукающем или в зычном трубном крике. Кроме того, молодые птицы принимают горизонтальную позу п втягивают шею совсем как взрослые, но вздергивают голову очень редко. Гораздо чаще они на мгновение вертикально вытягивают шего и тут же снова втягивают, держа голову при этом горизонтально, а не вздергивая ее. Таким образом, выпрашивающее движение взрослой чайки отличается от такого же движения молодой птицы полным отсутствием "насосного" элемента и несравненно более частым вздергиванием головы. Исходная позиция в обоих случаях одинакова, хотя молодые птицы, как правило, держат туловище более горизонтально.
Ту же позу, но в предельно горизонтальном положении (как у молодых птиц) принимает взрослая самка, не имеющая пары, а иногда и самцы, когда они "делают предложение", как описывалось выше.
Ритуальное кормление в брачном поведении
Таким образом, выпрашивание корма входит в систему полового поведения. И тут серебристая чайка не является исключением. Кормление и соответственно выпрашивание корма входят в ритуал ухаживания или предкопуляционного поведения у многих птиц и даже у некоторых других животных. Ценный обзор "ритуального кормления при ухаживании" у птиц был сделан Лэком [72]; сходные явления У других животных описал Майзепхаймер [82].
И здесь нам снова приходится рассматривать происхождение странного сочетания двух форм поведения, в данном случае опирающихся на побуждение кормить и сексуальное побуждение. Кормление, само собой разумеется, является одной из форм поведения родителей, которое определяется стремлением к продолжению рода и вступает в действие через довольно большой отрезок времени после того, как чисто половое поведение полностью прекратится. Забота о потомстве, как и половое поведение, зависит от степени стремления к продолжению рода. Нервные механизмы, лежащие в основе движений кормления, нормально действуют на последней стадии цикла размножения, которая паступает много позже стадии полового поведения. Поэтому движения кормления в системе полового поведения совершенно необоснованны.
Но "необоснованные" движения в половом поведении появляются столь же часто, как и в агрессивном, - факт, известный уже давно и постоянно подтверждающийся по мере того, как изучаются и сравниваются все новые виды животных. Как обычно, сравнение, показывая, что некоторые явления, кажущиеся на первый взгляд случайными, на самом деле распространены очень широко, привлекает внимание к проблемам, которые иначе могли бы остаться незамеченными.
В 1914 году Хаксли [57] описал "привычную чистку оперения" у чомг. Чистка оперения как часть ритуала ухаживания наблюдается у многих голубей, уток и шилоклювок. Сойки и некоторые райские птицы во время ухаживания ритуально чистят клювы. Самки, некоторых куриных птиц, когда самец производит перед ними ритуальную демонстрацию, делают клевательные движения. В малонаучных описаниях ухаживания у павлинов и индеек часто указывается, что самка "совершенно равнодушна" и клюет корм, словно самец ее вовсе не интересует. Я же убежден, что эти движения самки - явный признак сильного интереса, точно так же как чистка оперения у голубей свидетельствует об интенсивном стимулировании сексуального побуждения.
Тут, как и при изучении агрессивного поведения, полезно провести сравнение не только с птицами. У самца трехиглой колюшки с сильно стимулированным сексуальным побуждением мы наблюдаем элементы родительского поведения, хорошо нам известные по более позднему этану цикла размножения этой рыбки, - движения "обмахивания", которые обеспечивают аэрацию икры. Обмахивание - столь же обязательный компонент родительского поведения колюшки, как и насиживание у птиц. Когда на территории колюшки появляется самка, самец начинает интенсивно ухаживать за ней, но время от времени возвращается к гнезду и производит обмахивание, хотя там нет икры, которая нуждалась бы в аэрации.
Пытаясь объяснить подобные примеры, мы, естественно, склонны сравнивать их с другими проявлениями необоснованного поведения, о которых я говорил выше. Я указывал тогда, что необоснованные движения во время агрессивных стычек обеспечивают выход сильным и блокированным импульсам, поскольку стремление к бегству препятствует настоящему нападению. Таким образом, необоснованные движения - это не что иное, как смещенное поведение, возникающее из-за конфликта двигательных импульсов. Может быть, необоснованные движения во время ухаживания тоже объясняются не находящим исхода побуждением? Может быть, сексуальное побуждение, почему-то не преображающееся в соответствующие движения, находит выход через какие-то другие механизмы центральной нервной системы? Такой вывод, бесспорно, представляется наиболее правдоподобным. Доводы в его пользу вкратце сводятся к следующему. Одна из функций ухаживания заключается в стимулировании половых реакций у брачного партнера с помощью специфических сигналов. Эти реакции могут представлять собой немедленные действия или же "латентные" внутренние изменения, приводящие к постепенному возрастанию готовности к спариванию. Специфичность как сигналов, так и реакций партнера препятствует спариванию с особями другого вида. Обычно ухаживание бывает взаимным, то есть самка воздействует на самца в той же мере, что и самец на самку. Сигналы у самцов и у самок могут быть различными или, как это бывает у других видов, совершенно одинаковыми ("взаимное ухаживание").
Во многих случаях необоснованные движения проделываются в ситуациях, когда данное животное находится под воздействием сильных сексуальных импульсов, но не может совершить полового акта, так как от его партнера еще не поступило сигнала, стимулирующего этот акт. Птицы, а также многие другие животные в такой степени зависят от подобных раздражителей, что не предпримут никаких попыток к спариванию, если от партнера не поступило соответствующего стимула, разве только их внутреннее побуждение оказывается исключительно сильным. Для спаривания необходимы и внутреннее побуждение и поступающие от партнера стимулирующие раздражители. Ситуация, возникающая в момент ухаживания, и, главное, предкопуляционная ситуация характеризуются сильным внутренним сексуальным побуждением, но до тех лор, пока от партнера не поступило соответствующего сигнала, эти импульсы не находят выхода в половом акте. Поэтому, как и в агрессивной ситуации, здесь, хотя и по другой причине, внутреннее побуждение остается неудовлетворенным и, как и в агрессивной ситуации, необоснованные движения, по-видимому, представляют собой смещенное поведение.
Соответствующее поведение у колюшек
Гипотеза о происхождении "необоснованных" элементов ухаживания или предкопуляцпонного поведения, насколько мпе известно, не подвергалась экспериментальной проверке на птицах. Но в отношении рыб существуют очень интересные экспериментальные данные. Кроме того, сравнительное изучение предполагаемого смещенного поведения в процессе ухаживания у птиц показывает, что оно проявляется именно в те моменты, когда можно заметить признаки очень сильного, но не удовлетворенного сексуального побуждения. Дальнейшее исследование этих явлении было бы очень желательно по ряду причин. Но оио даст результаты только при условии систематических наблюдений за поведением и критической оценки обстоятельств, при которых возникают необоснованные движения. Изучение отдельных описаний в литературе или наблюдения от случая к случаю совершенно недостаточны для окончательных выводов.
Чрезвычайно интересно, что смещенное поведение, представляющее собой выход для сильного сексуального или агрессивного побуждения, воздействует и на партнера, который каким-то образом "понимает" его смысл. Когда смещенное поведение возникает из-за подавленного стремления к драке, противник пугается, когда же его порождает сильное сексуальное побуждение, оно стимулирует партнера к спариванию. Иначе почему, например, смещенное гнездостроительство у серебристых чаек вызывало бы реакцию не как гнездостроительство, а как угроза?
Разбирая движения угрозы, мы видели, что их угрожающий характер становится явным из-за наложения на них зачаточных движений драки. Что касается смещенного поведения в брачной ситуации, то мы еще не знаем, как возникла его стимулирующая функция. Нам пока известно лишь, что многие типы смещенного поведения, несомненно, имеют функцию стимулирующих раздражителей. Многие, по не все - и это привело к интересному открытию.
Ритуализация
Смещенное поведение как элемент ухаживания, воздействующий на партнера, и смещенное поведение как элемент угрожающего поведения, запугивающий противника, Довольно часто чуть-чуть отличаются от того поведения, которое стоит за ними. Настоящая чистка оперения у уток отличается от смещенной чистки, которая является элементом ухаживания. Последняя обычно ограничивается определенными движениями, посящими чисто формальный характер. Поэтому Селес [106] назвал такие движения "стилизованными", а Хаксли [58] предложил для них термин "ритуализовашше", которым я и буду пользоваться здесь.
Насколько нам известно, ритуализация, по-видимому, характерна для тех "заимствованных" движений, которые оказывают стимулирующее воздействие па поведение другой особи, а потому напрашивается вывод, что она представляет собой результат вторичного эволюционного процесса, который тесно связан с функцией стимулирования. Это становится еще более очевидным при сравнении разных типов такого ритуализованного смещенного поведения - ритуальность всегда делает движение более заметным. Во многих случаях это объясняется его "преувеличенностью". Смещенное поведение как раздражитель часто бывает преувеличенным, подчеркнутым. Той же цели служат яркие пятна на тех частях тела, которые легко демонстрируются при смещенном поведении. Отличпые примеры можно найти среди уток. Во время смещенной чистки оперения селезень кряквы поднимает крыло так, что становится видно красивое голубое* зеркальце на нем. У мандаринок смещенная чистка ограничивается одним ритуальным приглаживанием второстепенного махового пера, резко отличающегося от всех прочих: опахало у него не узкое и темно-зеленое, а очень широкое и ярко-оранжевое. Ритуализованным движением чистки самец словно старается обратить внимание самки на это перо. Совсем иная смещенная чистка оперения у селезня чирка-трескунка: он не заводит клюв за крыло, а только касается внешней его стороны там, где расположены голубые кроющие перья. Селезень пеганки добавил к ритуализованной чистке оперения звуковой аккомпанемент: быстро проводя клювом но стержням маховых перьев крыла, он производит низкий рокочущий звук.
*(Скорее фиолетовое. - Прим. ред. 114 )
Ритуализация не только делает движение более броским, но и придает ему особый характер, присущий лишь данному виду, что уменьшает возможность межвидового спаривания. Таким образом, сводя заимствованное движение к функции сигнала, процесс ритуализация тем самым затемняет его происхождение. Эволюция, возможпо, способствует развитию этой тенденции, так как заметное отличие заимствованного движения от его оригинала должно быть полезным для выживания вида. Я уже упоминал в главе 6, что форма угрожающего демонстративного поведения, называемая "кашлянием", очень похожа па движение, которое ее породило, а то и вовсе идентична ему. У серебристой чайки это движение( пока?) не ритуализовано. Но ритуализовано у смеющейся чайки, которая при смененном кашлянии взъерошивает перья. Большой интерес представляет "ошибка", которую на моих глазах допустил самец серебристой чайки, не распознавший угрозы в агивном кашлянии и принявший его за дружеское. Этот самец приблизился к самке на ее территории в отсутствие ее супруга. Самка встретила его кашлянием. (Поскольку она предварительно к нему не подошла, а также учитывая дальнейшее развитие событий, мы поняли, что кашляние было агрессивным.) Однако самец подбежал к пей и собрался встать рядом, как обычно делают супруги, но тут она поднялась и напала на него. Мне кажется, подобпые неувязки в системе общественных сигналов случались бы реже, если бы кашляние у серебристых чаек было ритуализовано, как у смеющейся чайки.
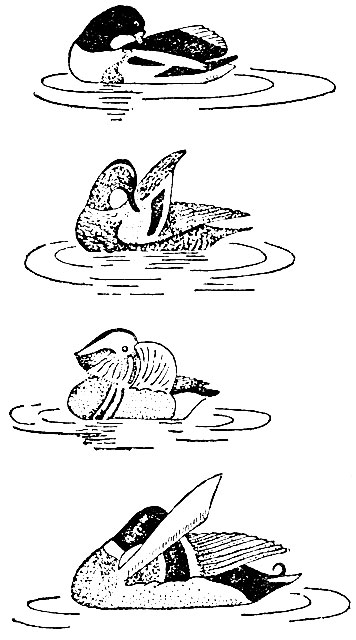
Рис. 15. Смещенная чистка оперения у ухаживающих селезней. Сверху вниз: пеганка, чирок-трескунок, мандаринка и кряква
Мы знаем, что родственные виды в сходных ситуациях часто демонстрируют одинаковое смещенное поведение. Мы наблюдали это у чаек и уток, но то же известно, например, о колюшках и журавлях. У различных видов в каждой из этих групп ритуализация развивалась по-разному, что привело к усилению специфичности, причем у каких-то видов она зашла так далеко, что в настоящий момент уже невозможно распознать происхождение столь ритуализованного смещенного поведения. В таких случаях мы были бы беспомощны, не будь у нас возможности провести сравнение между разными видами. Лоренц писал [77], что происхождение угрожающего демонстративного поведения у маньчжурского журавля было бы полностью затемнено, если бы сравнение не помогло нам распознать в нем крайне ритуализованную форму смещенной чистки оперения. Точно так же большинство наблюдателей, возможно, не сообразили бы, что прикасание к крылу у мандаринки - это тоже смещенная чистка оперения, не знай они менее ритуализованных ее форм у других видов.
Ритуализация имеет еще один интересный аспект, но о нем тут можно упомянуть лишь вкратце. Благодаря ритуализации движение все больше утрачивает сходство с породившим его движением. Отсюда следует, что происходят изменения в стоящих за этим механизмах центральной нервной системы. Другими словами, ритуализованное смещенное поведение приобретает все большую независимость, и все труднее решать, с каким побуждением оно связано - с инстинктом, у которого оно "позаимствовано", или с инстинктом, для выражения которого оно служит. Когда ритуализация заходит достаточно далеко, смещенное поведение утрачивает смещенность и включается в систему своего нового "хозяина".
Из-за относительной скудности материала все эти соображения остаются в известной мере предположительными.
Но, во всяком случае, очевидно, что изучение различных демонстративных форм поведения, связанных с угрожающим поведением и ухаживанием, могло бы дать очень многое. Проподя подробное сравнительное изучение родственных видов и стараясь в каждом случае по возможности выяснить на основе изложенной выше гипотезы происхождение демонстративных движений, мы могли бы лучше понять, каким образом и почему разные типы смещенного поведения стали такими, какими мы их видим теперь.
Вернемся, однако, к вздергиванию головы у серебристой чайки и проверим, можно ли в данном случае применить принципы, о которых шла речь выше. Поскольку вздергивание головы представляет собой существенную часть предкопуляциониого поведения и в то же время в нем легко распознается движение, которым молодые птицы выпрашивают корм, его следует рассматривать как смещенное поведение, дающее выход сексуальному побуждению. Далее, можно с уверенностью утверждать, что оно песет функцию раздражителя: вздергивание головы обычно вызывает у партнера ответную реакцию. Сравнение с исходным движением выпрашивания корма у молодых птиц убеждает нас, что оно ритуализовано очень мало, хотя выше я упоминал о некоторых различиях в форме движения и указал, что они могут объясняться определенной ритуализацией. Крик выпрашивания различается у взрослых и молодых птиц, но это, по-видимому, просто следствие общих изменений тембра голоса, которые обнаруживаются во всех криках в процессе индивидуального развития. Молодые птицы издают и крик тревоги, и трубный крик, и крик выпрашивания, но звучат они хрипло и пискливо, а не чисто и мелодично, как у взрослых. Следовательно, изменение голоса типично не только для крика выпрашивания. Морфологической структуры, сравнимой, скажем, с оранжевым пером мандаринки, тут нет. А потому ритуализация в данном случае, по-видимому, отсутствует.
Этот вывод согласуется с тем, что самка, выпрашивая корм, демонстрирует точно такое же поведение, как и тогда, когда она пытается побудить самца к спариванию. И для меня остается тайной, откуда самец знает, хочет ли самка есть или ею руководит сексуальное побуждение. Существуют, по-видимому, следующие возможности: либо самка в обоих случаях находится в одном п том же состоянии и любая альтернатива ее удовлетворяет, хотя и в разной степени, либо самец реагирует на какие-то мельчайшие различия, которые нам заметить не удалось.
Что касается первого предположения, не исключено, что вздергивание головы у самки всегда выражает сексуальное побуждение и что акт кормления его удовлетворяет, хотя и не в полной мере. На это указывает следующий факт: спаривание может произойти после кормления, обратного мы но наблюдали ни разу. Что касается второго предположения, то поскольку зрение чаек превосходит наше, они, вероятно, способны реагировать на сигналы, которые нам, несмотря на всю нашу опытность, не удается заметить - во всяком случае, пока мы пользуемся относительно грубыми методами, неизбежными при полевых наблюдениях.
Распознавание пола
Хотя вздергивание головы, по-видимому, бесспорно выражает активное сексуальное побуждение и заимствовано из движений выпрашивания корма, надо еще найти объяснение, почему для выхода сексуальных импульсов используется такая форма поведения, которая присуща неполовозрелым птицам. Как я уже говорил, это наблюдается у многих видов. Мне кажется, серебристая чайка может дать нам ключ к решению проблемы. Мы видели, что первым признаком появления сексуального побуждения у самца служит агрессивность по отношению к другим особям того же вида. Такая агрессивность направлена главным образом против самцов. Очень важно понять, что в этом заключается особая проблема. Чем объясняется такая избирательность? Почему самец не нападает и на самок? Иными словами, как чайки распознают пол своих сородичей? И тут нам опять помогает сравнение. Оно показывает, что у тех видов, которым присущ четкий половой диморфизм, самец распознает пол другой особи по оперению. Так, самец золотистого фазана начинает ухаживать за чучелом самки и нападает па чучело самца [86]. Если же самцы и самки данного вида имеют одинаковое оперение, самец распознает пол другой особи главным образом по поведению.
У серебристых чаек это, по-видимому, является правилом. Ответная реакция на первоначальную агрессивность самца у самок и у самцов совершенно различна. Самец либо обращается в бегство, либо сам начинает угрожать, а соответственно настроенная самка принимает описанную выше распластанную позу.
Распластанная поза во всех деталях представляет собой полную противоположность вертикальной угрожающей позе. Шея втянута, а не вытянута, что в случае драки ставит птицу в невыгодное положение; крылья не приподняты. Иными словами, самка не готовится к драке. Наоборот, она, как и вполне взрослые, но еще неполовозрелые молодые птицы, выражает покорность. И у выросших птенцов у самки причины для принятия такой позы одни и те же: им одинаково необходимо погасить потенциальную агрессивность, присущую самцам в брачный период. Очень интересно, что у молодых птнц эта поза появляется, только когда они утрачивают облик птенца и начинают выглядеть как взрослые чайки. Очевидно, пуховик достаточно отличается от взрослой птицы, чтобы не вызывать враждебной реакции; когда же птенец подрастает и делается похожим на взрослых птиц, у него, естественно, возникает необходимость в защитном поведении. Вскоре после окончания гнездового периода, когда птенцы становятся самостоятельными, тенденция вражды между особями берет верх над тенденцией терпимости по отношению к молодым птицам.
Покорность у самок
Идентичность распластанной позы самки с покорной позой, которая выработалась у вида для других конфликтных ситуаций между отдельными особями, проливает свет на уже упоминавшийся выше любопытный факт: хотя самки иногда бывают крупнее отдельных самцов, еще ни разу не сообщалось о том, чтобы самец образовал пару с самкой крупнее себя или же самка - с самцом мельче, чем она сама. Это лишний раз указывает на то, что. покорность самки представляет собой необходимое условие образования пары.
Агрессивность первой реакции самца на других птиц, как мне кажется, служит основной причиной тесной взаимосвязи между образованием пар и иерархическим доминированием. Первоначальная агрессивность самца привела к тому, что у самки должен был выработаться способ, позволяющий предотвратить нападение самца и вызвать у пего сексуальную реакцию. Для этой цели самка использует уже существующую форму поведения: принимает покорную позу, которая применяется во враждебных столкновениях совсем иного рода. Из-за агрессивности самца пары, в которых самка не подчинялась бы, попросту невозможны. Конечно, это относится только к видам с таким типом образования пар. Говоря, что самка использует уже существующую форму поведения, я вовсе не имею в виду, будто иерархические конфликты филогенетически старше сексуальных. Оба типа развивались одновременно, и, пожалуй, точнее будет сказать, что у данного вида выработалась определенная форма общественного поведения, которая должна предотвращать нападение в обоих типах конфликта.
Вполне возможно, что выпрашивание корма развилось из покорности самки как необходимого условия полового контакта с самцом. Если самке в процессе образования пары приходится принимать птенцовую позу, естественно, что этому будут сопутствовать и другие элементы птенцового поведения. Тому, что выпрашивание корма вошло в систему поведения при образовании пары, могли способствовать два обстоятельства. Во-первых, родительское побуждение у самца, достигающее полной силы только при появлении птенцов, может, как и все другие репродуктивные побуждения, в какой-то мере проявляться задолго до того, как в нем возникнет реальная необходимость. В результате на покорную позу самки он реагирует кормлением. Во-вторых, добавочный корм может быть полезен самке в период развития яиц в ее организме, а потому естественный отбор благоприятствовал закреплению супружеского кормления, едва оно возникло в зачаточной форме.
Вздергивание головы у самца
Но почему самец перед спариванием тоже демонстрирует поведение выпрашивания? Ему не нужно выражать покорность, и самка его никогда не кормит. А потому два указанных фактора, предположительно благоприятствовавшие развитию выпрашивания у самки, воздействовать на развитие такого поведения у самца никак не могли. Этому можно дать несколько объяснений, но ни для одного из них пет достаточно веских доказательств, так что пока лучше оставить вопрос открытым.
Не касаясь проблемы происхождения предкопуляционной формы поведения, вернемся к механизму этого поведения в его современном виде и зададимся вопросом, какие факторы прекращают вздергивание головы и позволяют перейти к собственно половому акту? Для самки - ато копуляционные действия самца. Она продолжает вздергивать голову до моментов спаривания. В промежутках между ними она вновь вздергивает голову. Это и понятно. Ведь если вздергивание головы представляет собой смещенное поведение, питаемое сексуальным побуждением, оно должно прекращаться сразу же, едва стоящее за ним побуждение найдет естественный выход.
С самцом дело обстоит иначе. В брачном поведении многих видов, как, например, у колюшек, смещенные действия продолжаются до тех нор, пока раздражитель, обеспечиваемый партнером, не стимулирует следующего этапа в цепи полового поведения. У серебристых чаек самец вспрыгивает на спину самки, не получив от нее повых сигналов: она лишь вздергивает и вздергивает голову, и никаких изменений в ее поведении как будто не происходит. Самец же постепенно вытягивает шею (движение намерения перед вспрыгиванием), все еще вздергивая голову, а затем внезапно вспрыгивает на спину подруги, одновременно переставая двигать головой и начиная испускать свой ритмичный брачный крик, по-видимому, тут возможно лишь одно объяснение: стимулирование накапливается, пока не достигается порог следующего звена в цепи брачного поведения самца. Мы уже знаем, какую важную роль в стимулировании сексуального побуждения самца играет вздергивание головы у самки, а потому такой вывод представляется более правдоподобным, чем предположение, что самец, вздергивая голову, сам стимулирует себя.
Мне кажется, этот краткий анализ дает нам некоторое представление о том, как могла возникнуть подобная сложная форма совместного поведения. В основе ее, по-видимому, лежит двойственность поведения самца. Драки весной, как мы убедились, необходимы, чтобы обеспечить пространство, нужное для нормального существования пары и ее потомства. Побуждения к драке у самца должны каким-то образом подавляться самкой, иначе продолжение рода окажется невозможным. Единственный действенный способ - принять позу покорности. Но подавить агрессивность самца еще мало, необходимо стимулировать его сексуальное побуждение.
Это достигается благодаря смещенному поведению, которое возникает из-за неудовлетворенности сексуального побуждения самки, поскольку вначале такое побуждение у самца отсутствует. Смещенное поведение, то есть смещенное выпрашивание корма самкой, включается в систему потому, что вызывает у самца сексуальную реакцию. Таким образом, сложная форма взаимного поведения возникает опять-таки благодаря наличию по меньшей мере двух побуждений, которые в гнездовой период постоянно находятся в активном состоянии. У такого поведения есть и добавочная функция: животные обладают врожденными избирательными реакциями на раздражители, присущие только их виду, а это препятствует спариванию с особями другого вида. Наличие двух побуждений обеспечивает сложность брачного поведения. Виды с одинаковым брачным поведением смешивались бы в один вид - вот почему форма брачного поведения у существующих в настоящее время видов различна и действует как репродуктивно изолирующий механизм.
Скрещивание с клушами
В этой связи интересно заметить, что одна из ближайших родственниц западноевропейской серебристой чайки, клуша, несмотря на явное различие в окраске оперения и заметную разницу в голосе, иногда спаривается с серебристой чайкой. Это означает, что формы брачного поведения у них еще пе настолько дифференцировались, чтобы полностью исключить их скрещивание в естественных условиях. На острове Терсхеллинг такие смешанные пары наблюдались с 1928 года (см. фото 15). Потомство этих пар также способно давать потомство - со времени появления клуш и в других колониях серебристой чайки мы не раз наблюдали, как птенцов выводили явные гибриды этих двух видов. В 1949 году я заснял на кинопленку гибрида-самца с его подругой, чистокровной серебристой чайкой, и птенцами в пуховом наряде. Мы предполагали в следующем году поймать эту пару и вырастить их потомство в неволе, но в 1950 году гибрид в колонии не появился.
Однако массового скрещивания между этими двумя видами не происходит; как правило, все исчерпывается одной смешанной парой. В 1928 году на Терсхеллинге было четыре пары чистокровных клуш и одна смешанная пара. В послевоенные годы там обычно насчитывалось более двадцати чистых пар и опять же всего одна смешанная, а то и вовсе ни одной. В Вассенаре в 1949 году было пять пар клуш и та самая пара, которую я снимал. Поэтому представляется вероятным, что смешанные пары образуются, только когда клуша селится в колонии, где для нее нет партнера ее вида. Если это так, то гибридизация должна быть типичной лишь для популяций, осваивающих новые места, причем ее вероятность уменьшается по мерс увеличения общего числа клуш, гнездящихся в новой колонии.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ornithology.su/ 'Орнитология'